Supplement review: Nicotinamide riboside
Nicotinamide riboside (or NR for short) is a form of vitamin B3. So, before delving into the depths of NR, let us first take a short look at vitamin B3 in general.
Our history starts in 1735 when the Spanish doctor Gasper Casal described a disease in poor farmers characterised by sun-sensitive Dermatitis (inflammation of the skin), Diarrhea, Dementia and eventually Death (the four ‘Ds’). This same disease became an epidemic in the southern states of the US around the turn of the 20th century. It is estimated that at least 100,000 Americans died from this disease between 1907 and 1940. This disease was pellagra and is common in people whose diet consists nearly completely out of corn and are low in meat. While the role of diet in the development of pellagra was already suspected by Casal and the French had successfully eradicated pellagra by limiting the cultivation of corn many continued to believe that pellagra was an infectious disease. The American physician and epidemiologist Joseph Goldberger definitively proved that pellagra was caused by a deficient diet in the early 20th century after conducting dietary experiments on inmates. When the inmates were placed on a corn-rich diet with little other food sources they started to develop pellagra. Eventually researchers were able to figure out that pellagra was caused by a deficiency of two nutrients vitamin B3 and tryptophan (see below to find out the role of tryptophan) (Wan et al., 2011; Yang and Sauve, 2016).
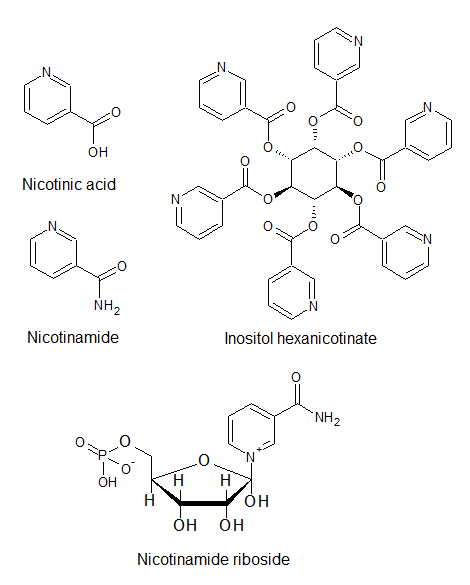
There are several different forms of vitamin B3 which all have slightly different properties but share the fact that the body can use them equally to produce the vitamin B3 analogs that we need to stay alive. Nicotinic acid is a form of vitamin B3 that in high dosages is used as a drug to lower blood lipids but its use has been mostly replaced by statin drugs, although recently there’s a resurgence of interest in nicotinic acid due to its wide effect on many blood lipids. Of all drugs used to lower blood lipids nicotinic acid remains unique in the fact that it positively influences a large array of blood lipids (it lowers LDL, Lp(a) and triglycerides while increasing HDL) hence it is sometimes called a “broad-spectrum lipid drug”. Interestingly, nicotinamide, while very similar to nicotinic acid in chemical structure (see image 1), does not have blood lipid lowering properties (Ballantyne, 2009; Carlson, 2005; Jacobson et al., 2012). A major problem with the use of high dose nicotinic acid therapy is that it causes flushing leading to low compliance. Hence extended-release forms that cause less severe flushing have been developed. In shops one may also find “flush-free” niacin, this is inositol hexanicotinate but this molecule does not improve lipid levels. Researchers are working on the development of newer non-flush nicotinic acid derivatives against cardiovascular disease including transdermal (through the skin) delivery (Jacobson et al., 2012). NR is a form of vitamin B3 found in low amounts in milk and yeast products (Chi and Sauve, 2013) and NR together with pterostilbene are the major components of “Basic”, the supplement produced by the biotech company Elysium Health. NR is a very recent supplement as the production of this molecule remained difficult until recently. This also made studies with NR difficult. In July 2013 the first NR supplement, NIAGEN, hit the market (Chi and Sauve, 2013). Today NR is available as capsules or as bulk powder from many supplement companies.
Image 1: Different forms of vitamin B3. Image generated using ACD/ChemSketch®.
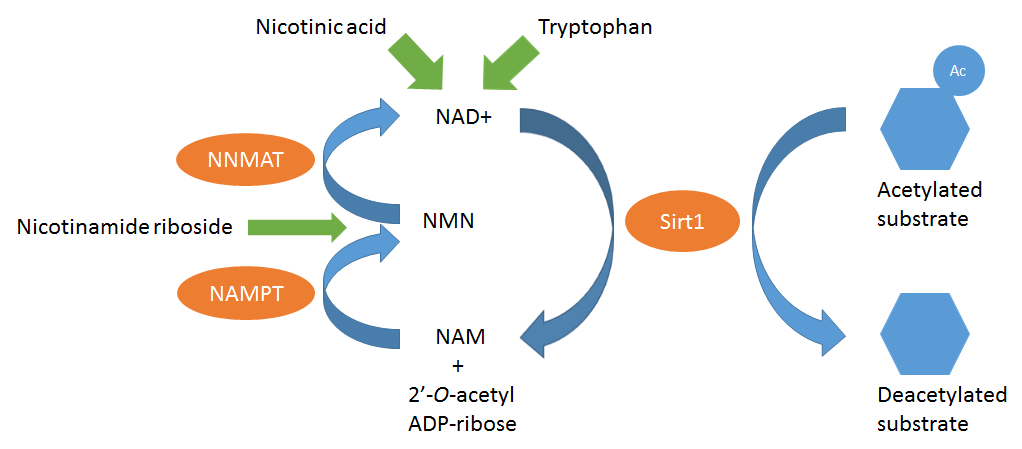
Our bodies need vitamin B3 as precursor for NAD+, an electron transfer molecule that has a crucial role to play in energy production. In addition NAD+ is needed in certain signaling pathways and is also converted into NADPH which is involved in detoxification of reactive oxygen species (ROS). The amino acid tryptophan can also be converted into NAD+ explaining why tryptophan deficiency can cause pellagra (image 2).
NR appears to be very safe with a safety profile similar to that of nicotinamide. No adverse effects were observed in rats for doses below 300 mg/kg/day (Conze et al., 2016). Supplement companies typically advice 100-300mg a day.
Recently, several animal studies have been published that show the effectiveness of NR against several diseases. Supplementation of mice with NR prevented weight gain in animals fed a high fat diet despite the fact that NR supplemented animals tended to eat more. Blood sugar levels were similar in NR supplemented animals compared to control animals but insulin levels are lower indicating increased insulin sensitivity in the NR supplemented animals. The researchers next performed a battery of standard diabetes tests and found that the NR-supplemented animals performed better compared to the control animals (Cantó et al., 2012). Similarly, Charles Brenner and colleagues treated pre-diabetic and diabetic mice with NR and found improvements in glucose tolerance, reduced weight gain, improved protection against liver damage from diabetes and protection against diabetic neuropathy (nerve damage caused by diabetes) (Trammell et al., 2016b). Levels of amyloid-beta were decreased and cognitive function was improved in a mouse-model of Alzheimer’s disease treated with NR for three months (Gong et al., 2013). NR supplementation improved mitochondrial function in a mouse model of mitochondrial disease (Cerutti et al., 2014). NR also protected mice from noise-induced hearing loss (Brown et al., 2014) and improves liver regeneration in mice (Mukherjee et al., 2016). Johan Auwerx and colleagues demonstrated that NR supplementation delayed disease progression in a mice model of mitochondrial myopathy (muscle disease) (Khan et al., 2014). NR supplementation improved muscle function and heart problems in a mouse model of muscular dystrophy (Ryu et al., 2016). Furthermore, NR supplementation reversed muscle fiber degeneration in mice missing the Nampt enzyme involved in NAD+ recycling (see image 2) (Frederick et al., 2016). However, it should be pointed out that one study found that NR supplementation decreased exercise performance in normal rats by 35% (Kourtzidis et al., 2016).
In 2007 it was discovered that NR increases replicative lifespan in yeast (Belenky et al., 2007). Replicative lifespan is the number of times that a cell, referred to as the mother cell, can bud of daughter cells before it permanently stops dividing. Uptake of endogenous NR is also needed for calorie restriction-induced lifespan extension (Lu et al., 2009). More recently, it was shown that NR supplementation is able to slightly increase the lifespan of mice (Zhang et al., 2016). Mean lifespan was increased by 4.7% and maximum lifespan was also increased (as illustrated by the graph in the paper) but the authors did not calculate the statistical significance for the maximum lifespan data (Zhang et al., 2016). Supplementation of the roundworm C. elegans with NAD+ (Hashimoto et al., 2010), nicotinamide or 1-methylnicotinamide (Schmeisser et al., 2013) has also been found to increase lifespan.
While NR supplementation seems to have multiple benefits in laboratory animals we should notice that research in this area has only started. To the best of our knowledge no human data on the health effects of NR has been published so far. Also the reduced exercise performance in rats fed NR is a warning that we shouldn’t implement NR supplementation in healthy humans as of yet (Kourtzidis et al., 2016). In a few years more data about NR should be available allowing us to make a much more informed decision.
Sirt1 is a well-known enzyme that removes a chemical group known as an acetyl group from proteins, most notably from histones (the proteins around which DNA is wrapped). By chemically modifying the histone proteins, Sirt1 is able to regulate the expression of genes. Multiple studies have found that overexpression of Sirt1 (or its homologs) increases lifespan in yeast, fruit flies, and the roundworm C. elegans. However, a 2011 Nature paper found that the lifespan extending effects in fruit flies and C. elegans were caused by unrelated background mutations (Burnett et al., 2011). Sirt1 is also famous as the reputed target of the geroprotector resveratrol. However, multiple studies have found that resveratrol does not bind Sirt1 (Calamini et al., 2010). In the original Sirt1 studies a fluorescent peptide was used to measure the Sirt1 activity and this peptide induced a binding of resveratrol to Sirt1 while no such binding takes place in the absence of this ‘artificial measuring tool’. However resveratrol may also work through other mechanisms such as mTOR inhibition (Park et al., 2016) or inhibition of phosphodiesterases leading to AMPK activation (Park et al., 2012).
Yang et al. (2007) has shown that NR increases NAD+ levels by up to 270% in cultured mammalian cells. Interestingly, a recent human study showed that oral administration of 1,000 mg NR increases human blood NAD+ levels by 270% (Trammell et al., 2016a). Johan Auwerx and colleagues have demonstrated that administration of NR to mice increased NAD+ concentrations and activates Sirt1 (Cantó et al., 2012). Sirt1 needs NAD+ to dump the acetyl group on that it removes from its targets. Hence without NAD+, Sirt1 cannot do its enzymatic function. In the process the NAD+ molecule splits in two parts, one of which is nicotinamide (NAM). As you might remember nicotinamide is one form of vitamin B3. Nicotinamide can then be converted back in NAD+. Because nicotinamide is the end product of the reaction catalysed by Sirt1 it is expected that the presence of nicotinamide will inhibit Sirt1 activity. Indeed, nicotinamide is a well known inhibitor of Sirt1. In addition to Sirt1, NR also activates Sirt3 and mitochondrial biogenesis (Brown et al., 2014; Cantó et al., 2012; Gong et al., 2013).
Image 2: Sirt1 removes acetyl groups (round circle) from substrates (such as proteins). On the left is the circular mechanism in which NAD+ is regenerated from the end products of the Sirt1-catalysed reaction. In green are the two ways in which new molecules can enter in this cycle. Nicotinic acid and tryptophan are direct precursors of NAD+ while nicotinamide riboside enters this cycle by conversion into NMN.
Overexpression of NAMPT, an enzyme involved in recycling the end products from the Sirt1 reaction back to NAD+, extends replicative lifespan of human cells in culture (van der Veer et al., 2007; Ho et al., 2009). Restricting glucose in the medium used to grow cells in leads to an upregulation of NAMPT and increased NAD+ levels leading to an increase in replicative lifespan (Yang et al., 2015). Furthermore, overexpression of NAMPT in human endothelial cells (cells that line the blood vessels) is able to protect these cells from the harmful consequences of high glucose levels (Borradaile and Pickering, 2009). Similar results have been obtained in yeast. Yeast has an enzyme called Pnc1 that converts nicotinamide into nicotinic acid and thereby removes the inhibitory effect of nicotinamide on Sir2 (the yeast equivalent of Sirt1). Indeed, overexpression of Pnc1 was found to stimulate Sir2 activity (Gallo et al., 2004).
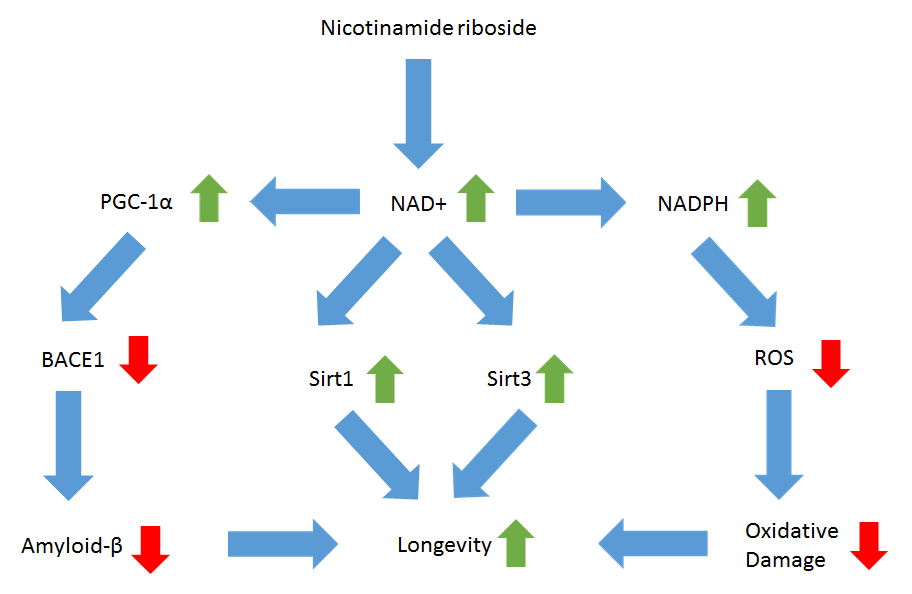
Image 3: Mechanisms by which NR promotes longevity. The color of the arrows refers to decreases (red) or increases (green) and does not reflect if these changes or beneficial or harmful.
References
Ballantyne CM (ed.) (2009). Clinical Lipidology: A Companion to Braunwald's Heart Disease, 1st edition. Saunders Elsevier, Philadelphia.
Belenky P et al. (2007). Nicotinamide riboside promotes Sir2 silencing and extends lifespan via Nrk and Urh1/Pnp1/Meu1 pathways to NAD+. Cell 129(3): 473-484.
Borradaile NM, and Pickering JG (2009). Nicotinamide phosphoribosyltransferase imparts human endothelial cells with extended replicative lifespan and enhanced angiogenic capacity in a high glucose environment. Aging Cell 8: 100-112.
Brown KD et al. (2014). Activation of SIRT3 by the NAD⁺ precursor nicotinamide riboside protects from noise-induced hearing loss. Cell Metab 20(6): 1059-1068.
Burnett C et al. (2011). Absence of effects of Sir2 overexpression on lifespan
in C. elegans and Drosophila. Nature 477: 482-485.
Calamini B et al. (2010). Pleiotropic mechanisms facilitated by resveratrol and its metabolites. Biochemical J 429(2): 273-282.
Cantó C et al. (2012). The NAD(+) precursor nicotinamide riboside enhances oxidative metabolism and protects against high-fat diet-induced obesity. Cell Metab 15(6): 838-847.
Carlson LA (2005). Nicotinic acid: the broad-spectrum lipid drug: a 50th anniversary review. J Intern Med 258(2): 94-114.
Cerutti R et al. (2014). NAD(+)-dependent activation of Sirt1 corrects the phenotype in a mouse model of mitochondrial disease. Cell Metab 19(6): 1042-1049.
Chi Y, Sauve AA (2013). Nicotinamide riboside, a trace nutrient in foods, is a vitamin B3 with effects on energy metabolism and neuroprotection. Curr Opin Clin Nutr Metab Care 16(6): 657-661.
Conze DB et al. (2016). Safety assessment of nicotinamide riboside, a form of vitamin B3. Hum Exp Toxicol [Epub ahead of print].
Frederick DW et al. (2016). Loss of NAD homeostasis leads to progressive and reversible degeneration of skeletal muscle. Cell Metab 24(2): 269-282.
Gong B et al. (2013). Nicotinamide riboside restores cognition through an upregulation of proliferator-activated receptor-γ coactivator 1α regulated β-secretase 1 degradation and mitochondrial gene expression in Alzheimer's mouse models. Neurobiol Aging 34(6): 1581-1588.
Hashimoto T et al. (2010). Nicotinamide adenine dinucleotide extends the lifespan of Caenorhabditis elegans mediated by sir-2.1 and daf-16. Biogerontology 11(1): 31-43.
Ho C et al. (2009). SIRT1 markedly extends replicative lifespan if the NAD+ salvage pathway is enhanced. FEBS Lett 583(18): 3081-3085.
Jacobson EL et al. (2012). Niacin: Vitamin and antidyslipidemic drug. In: Stranger O (ed.). Water soluble vitamins: Clinical research and future application. Subcellular Biochemistry 56. Springer, Dordrecht.
Khan NA et al. (2014). Effective treatment of mitochondrial myopathy by nicotinamide riboside, a vitamin B3. EMBO Mol Med 6(6): 721-731.
Kourtzidis IA et al. (2016). The NAD(+) precursor nicotinamide riboside decreases exercise performance in rats. J Int Soc Sports Nutr 13: 32.
Lu SP et al. (2009). Assimilation of endogenous nicotinamide riboside is essential for calorie restriction-mediated life span extension in Saccharomyces cerevisiae. J Biol Chem 284(25): 17110-17119.
Mukherjee S et al. (2016). Nicotinamide adenine dinucleotide biosynthesis promotes liver regeneration. Hepatology [Epub Ahead of Print].
Park SJ et al. (2012). Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting cAMP phosphodiesterases. Cell 148(3): 421-433.
Park D et al. (2016). Resveratrol induces autophagy by directly inhibiting mTOR through ATP competition. Scientific Reports 6: 21772.
Schmeisser K et al. (2013). Role of sirtuins in lifespan regulation is linked to methylation of nicotinamide. Nat Chem Biol 9(11): 693-700.
Trammell SAJ et al. (2016a). Nicotinamide riboside is uniquely and orally bioavailable in mice and humans. Nature Communications 7: 12948.
Trammell SA et al. (2016b). Nicotinamide Riboside Opposes Type 2 Diabetes and Neuropathy in Mice. Sci Rep 6: 26933.
van der Veer E et al. (2007). Extension of human cell lifespan by nicotinamide phosphoribosyltransferase. J Biol Chem 282: 10841-10845.
Wan P et al. (2011). Pellagra: a review with emphasis on photosensitivity. Br J Dermatol 164: 1188-1200.
Yang T et al. (2007). Syntheses of nicotinamide riboside and derivatives: effective agents for increasing nicotinamide adenine dinucleotide concentrations in mammalian cells. J Med Chem 50(26): 6458-6461.
Yang NC et al. (2015). Up-regulation of nicotinamide phosphoribosyltransferase and increase of NAD+ levels by glucose restriction extend replicative lifespan of human fibroblast Hs68 cells. Biogerontology 16(1): 31-42.
Yang Y, Sauve AA (2016). NAD+ metabolism: Bioenergetics, signaling and manipulation for therapy. Biochim Biophys Acta 1864(12): 1787-1800.
Zhang H et al. (2016). NAD⁺ repletion improves mitochondrial and stem cell function and enhances life span in mice. Science 352(6292): 1436-1443.