Abstract
Rapamycin (Rap) and its derivatives, called rapalogs, are being explored in clinical trials targeting
cancer and neurodegeneration. The underlying mechanisms of Rap actions, however,
are not well understood. Mechanistic target of rapamycin (mTOR), a lysosomelocalized
protein kinase that acts as a critical regulator of cellular growth, is believed to mediate
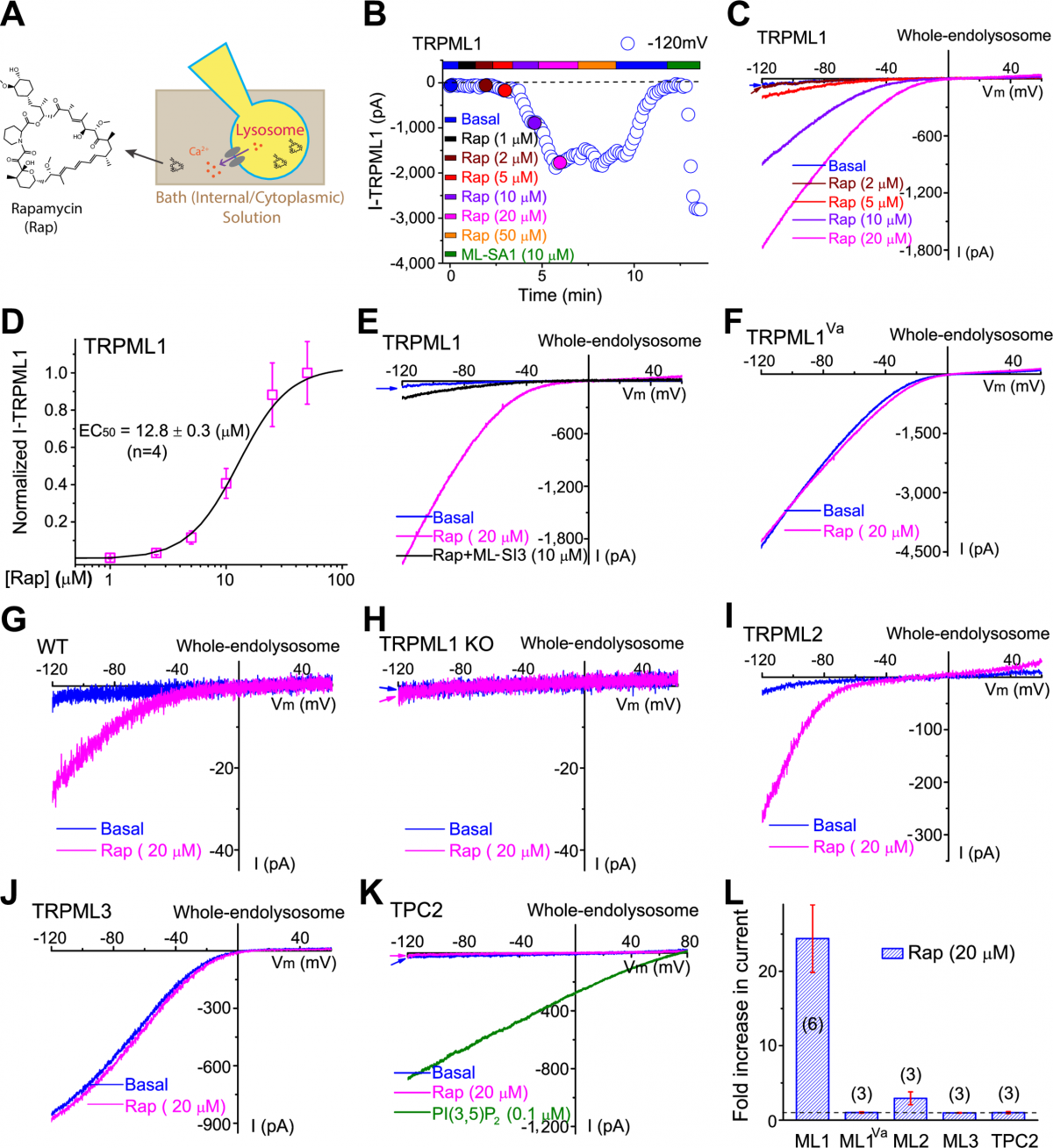
most Rap actions. Here, we identified mucolipin 1 (transient receptor potential channel
mucolipin 1 [TRPML1], also known as MCOLN1), the principle Ca2+ release channel in the
lysosome, as another direct target of Rap. Patch-clamping of isolated lysosomal membranes
showed that micromolar concentrations of Rap and some rapalogs activated lysosomal
TRPML1 directly and specifically. Pharmacological inhibition or genetic inactivation
of mTOR failed to mimic the Rap effect. In vitro binding assays revealed that Rap bound
directly to purified TRPML1 proteins with a micromolar affinity. In both healthy and disease
human fibroblasts, Rap and rapalogs induced autophagic flux via nuclear translocation of
transcription factor EB (TFEB). However, such effects were abolished in TRPML1-deficient
cells or by TRPML1 inhibitors. Hence, Rap and rapalogs promote autophagy via a
TRPML1-dependent mechanism. Given the demonstrated roles of TRPML1 and TFEB in
cellular clearance, we propose that lysosomal TRPML1 may contribute a significant portion
to the in vivo neuroprotective and anti-aging effects of Rap via an augmentation of autophagy
and lysosomal biogenesis.
Introduction
Rapamycin (Rap) is a natural macrocyclic compound that was initially isolated from Streptomyces
hygroscopicus as an antifungal agent [1]. Because Rap was shown to have robust immunosuppressive
and antiproliferative efficacy [2], Rap derivatives (rapalogs; see S1 Fig) with
improved pharmacokinetic properties have been developed in the industry, including temsirolimus
(Tem), everolimus (Eve), deforolimus (Defo), zotarolimus (Zota), WYE-592, and ILS-
920 [3, 4]. Since 1999, Rap (brand name Sirolimus) and several rapalogs have been approved
by the United States Food and Drug Administration for clinical trials testing their ability to
target cancer cells and to alleviate metabolic and neurodegenerative diseases [3, 4]. More
recently, Rap was also shown to extend life span across diverse organisms ranging from flies to
mammals [4, 5]. Hence, elucidating the molecular mechanisms of Rap bioactivities is of great
value for both basic and clinical research.
The first identified target protein of Rap was discovered in yeast and named target of rapamycin
(TOR) [6, 7]. TOR, now renamed mechanistic target of rapamycin (mTOR), is a serine
and/or threonine kinase that is highly conserved in eukaryotes [6, 7]. Although multiple cellular
locations have been reported, there is now a consensus that mTOR is localized predominantly
on the membranes of lysosomes under nutrient-rich conditions [8]. In response to environmental
changes, such as nutrient availability, mTOR kinase activity is switched on and off through
the formation of alternate protein complexes—mTOR complex 1 (mTORC1) and mTORC2—
and through association with and dissociation from lysosomal membranes. Known mTOR substrates
include, but are not limited to, UNC-5–like autophagy activating kinase (ULK1; also
known as autophagy-related protein 1 homolog), p70 ribosomal protein S6 kinase (S6K), 4E
binding protein 1 (4E-BP1), and transcription factor EB (TFEB) [9]. Rap acts as a high-affinity
(nM range) allosteric inhibitor of mTORC1 (hereafter referred to as mTOR) that blocks mTOR
substrate recruitment by binding to the FK506 binding protein (FKBP) and the rapamycin binding
(FRB) domain of mTOR, forming a ternary FKBP12-Rap-mTOR complex [3, 4].
Both the anticancer and immunosuppressive effects of Rap are likely due to its inhibition of
cell proliferation via mTOR, which integrates a number of signaling pathways in the cell and
has thus emerged as a major regulator of cellular proliferation and growth [7]. However,
mTOR inhibition also induces autophagy, a lysosome-dependent cellular survival mechanism
that supplies recycled nutrients by degrading obsolete cellular components [10]. Defective
autophagy may hasten aging and enable the pathogenesis of numerous diseases, including cancer
and neurodegenerative diseases [4]. Hence, autophagy induction caused by mTOR inhibition
may also explain many of the reported effects of Rap, especially neuroprotection and antiaging
effects [2, 11].
The basic autophagic process consists of autophagosome formation, autophagosome–lysosome
fusion, and lysosomal degradation [12]. Nutrient insufficiency is a potent inducer of
autophagy, in which the loss of nutrients (e.g., amino acids) causes mTOR inhibition. Subsequently,
dephosphorylation of ULK1, a major mTOR target, primes phagophore initiation
[12]. Rap can mimic the effect of starvation on ULK1-mediated autophagy induction [12].
Although all rapalogs inhibit mTOR potently, their clinical efficacies vary [13]. Rapalogs with
relatively low mTOR binding affinities (e.g., WYE-592 and ILS-920) exhibit neuroprotective
effects at least as potent as that of their counterparts with higher mTOR binding affinities [3].
Furthermore, although mTOR is inhibited much more potently by its catalytic inhibitors (e.g.
Torin-1), in vivo beneficial effects have not been observed for these potent inhibitors [14].
Hence, Rap may have other targets besides mTOR in the autophagy pathway.
Sustained autophagy requires lysosome activation, reformation, and biogenesis [12, 15, 16].
Under conditions when lysosome function is compromised, such as in neurodegenerative
Lysosomal Ca2+ channel is adiseases and lysosome storage diseases (LSDs), it is unlikely that an increase in autophagosome
formation alone could produce beneficial effects related to cellular clearance. Nutrient starvation,
a physiological inducer of autophagy, promotes both autophagosome formation and lysosome
biogenesis. Upon starvation-induced mTOR inhibition, TFEB, a key regulator of
autophagy and lysosome biogenesis [17], undergoes rapid activation via dephosphorylation
and cytosol-to-nucleus translocation [17–20]. Starvation may also activate mucolipin 1
(MCOLN1; also known as transient receptor potential channel mucolipin 1 [TRPML1]), a
lysosomal Ca2+ channel required for TFEB activation via the Ca2+-dependent phosphatase calcineurin
[21, 22]. Activation of TFEB, in turn, up-regulates TRPML1 expression [23]. Therefore,
TRPML1 and TFEB may constitute a positive-feedback loop that boosts lysosomal
biogenesis and autophagy under lysosomal stress conditions. Indeed, up-regulation of either
TFEB or TRPML1 has been reported to benefit several LSDs, including Pompe disease and
Niemann-Pick type C (NPC) disease, as well as common neurodegenerative diseases, including
Alzheimer disease [15, 17, 24, 25].
In the present study, we found that the TRPML1-TFEB-autophagy pathway is directly activated
by Rap and some rapalogs. Employing biomolecular interaction assays and whole-endolysosome
electrophysiology, we demonstrated that Rap bound directly to TRPML1 and
specifically activated TRPML1 independent of mTOR.
..../....