
































.
F U L L A C C E S S S O U R C E : ResearchGate
ABSTRACT
The T cell compartment undergoes characteristic changes with age, which contribute to increased incidence andseverity of infections and reduced immunogenicity and efficacy of many vaccines in the older population.Production of naïve T cells is severely impaired due to a decreased output of lymphoid cells from the bonemarrow and the involution of the thymus. At the same time, antigen-experienced, highly differentiated T cellsaccumulate resulting in a diminished T cell receptor repertoire. These cells show some similarities with senescentcells, such as shorter telomers, accumulated DNA damage and metabolic changes. Latent infection withCytomegalovirus also impacts the T cell compartment and aggravates several of its age-associated changes. Lossof CD28 expression is one hallmark of T cells after repeated antigenic stimulation, but CD28-T cells cannot beconsidered truly senescent as e.g. they are still able to proliferate upon adequate stimulation. Several additionalmarkers have been suggested in order to define a potential fully senescent T cell population, but no consensusdefinition has been reached so far. It has been postulated that highly differentiated senescent-like T cells areunable to eliminate other senescent cell types. Removal of senescent non-immune cells has been shown to bebeneficial for the organism and a reliable definition of senescent T cells is essential for an extension of this concept to T cells.
1. Introduction
Incidence and severity of many infectious diseases is increased inthe older population (Gavazzi and Krause, 2002;Kline and Bowdish,2016). There are many causes for this phenomenon including anato-mical and physiological changes and an increased risk due to hospita-lization or invasive procedures, but importantly, age-related changes inthe functionality of the immune system, summarized as im-munosenescence also play an important role. Although im-munosenescence affects both the innate and adaptive immune system,these changes are more pronounced in adaptive immune cells(Montecino-Rodriguez et al., 2013).
In this review, we summarize age related changes of T cells anddescribe the link between antigen-experienced, highly differentiated Tcells and cellular senescence. Finally, the intriguing possibility that“classical”cellular senescence may be supported by the aging of theimmune system is discussed.
2. Age-related changes in T cell development, function and repertoire
T cell precursors originate within the bone marrow (BM). Multipotent hematopoietic stem cells (HSCs) residing in the BM dif-ferentiate into common lymphoid progenitors (CLPs), giving rise to Band T lymphocytes and natural killer (NK) cells, or into commonmyeloid progenitors (CMPs), which can differentiate into granulocytes,monocytes, erythrocytes, and platelets. In older adults, HSCs lose theirself-renewal capacity (De Haan and Lazare, 2018). In addition, pre-ferential myeloid commitment in old age has been documented (Panget al., 2017;Wahlestedt et al., 2017). Specifically, increased frequenciesof myeloid cells in peripheral blood (PB), at both steady state and upontransplantation, have been observed. In parallel, preferential myeloiddifferentiation is associated with reduced lymphoid output of HSCs(Beerman et al., 2010;Young et al., 2016). In order to differentiate intonaïve T cells, precursors need to migrate to the thymus, the centrallymphoid organ responsible for the development and production ofmature naïve T cells (Koch and Radtke, 2011). T cell maturation iscontrolled by positive and negative selection, which ensure reactivityand removal of non-functional or autoreactive cells. After this process,mature T cells are released into the circulation and continuously re-circulate between blood, lymph, lymphoid organs and tissues.
The broad spectrum of changes affecting the immune system in oldage are known as immunosenescence, This process affects the adaptiveand, to a lesser extent, the innate immune system and is known to n suggested (Xu andLarbi, 2017). We will discuss the phenomenon of senescence in thecontext of T cells in more detail below.Recently, a link was found between decreased T cell output due tophysiological thymic involution and age-dependent accumulation ofhighly differentiated T cells. Specifically, reduced maturation of newnaïve T cells may lead to accelerated T cell homeostatic proliferation ofalready existing naïve T cells, supporting the generation of late differ-entiated T cells as a result (Sato et al., 2017). Alternatively, repeatedexposure to antigens (i.e. chronic viruses) may support the accumula-tion of this cell type.Similar to other cell types, DNA damage accumulates in T cells withage. Although DNA damage in naïve T cells remains low even in olderadults, it increases in memory T cells from healthy aged individuals(Park and Larsson, 2011). The consequences of damage to mitochon-drial DNA, a typical hallmark of aging, have not yet been investigatedin detail. The process of immunosenescence is accelerated in patientswith rheumatoid arthritis (RA). T cells show features of prematureaging, with shortened and damaged telomeres, increased production ofpro-inflammatory cytokines and impaired DNA repair systems (Goronzyand Weyand, 2012;Weyand et al., 2014). In particular, pro-in-flammatory properties of T cells in RA patients are connected to in-efficiency of DNA repair and telomere structural instability. In com-parison to T cells from healthy age-matched donors, RA T cells generateless ATP, because of a repression of the glycolytic enzyme phospho-fructokinase (PFKFB3), and produce high amounts of NADPH, due tothe overexpression of glucose-6-phospate dehydrogenase (G6PD), therate-limiting enzyme of the pentose phosphate pathway (Yang et al.,2013, 2016). The disbalance between ATP and NADPH generationsupports the consumption of reactive oxygen species (ROS), leading to a surplus of reductive elements, which directly promotes a Th1 and Th17commitment (Shen et al., 2017). In addition, metabolic reprogrammingin RA T cells has been linked to the DNA repair system. The DNA repairand cell cycle kinase Ataxia telangiectasia mutated (ATM) is a redoxsensor activated by ROS. In T cells from RA patients, ATM activity isreduced, therefore leaving such cells with a defective DNA repairsystem and allowing the cells to bypass the G2/M cell cycle checkpoint(Weyand and Goronzy, 2017;Yang et al., 2016).

In mice, defects in chaperone-mediated autophagy can be observedin T cells during aging (Valdor et al., 2014). In addition, decreasedlevels of basal macroautophagy have been observed in human CD8+Tcells (Phadwal et al., 2012). Increased expression of dual specificphosphatase 4 (DUSP4), which terminates nuclear extracellular-signalregulated kinase (ERK) signaling and impairs proliferation and effectorfunctions, has been documented in old CD4+memory T cells (Yu et al.,2012). This is the result of increased activation of the metabolic masterregulator AMP-activated protein kinase (AMPK) taking place inmemory T cells themselves. The importance of AMPK in controllingimmune stimuli and T cell metabolic fitness has been reported, there-fore that this kinase may function as a metabolic sensor for T cells(Lanna et al., 2014;Ma et al., 2017). In senescent-like CD45RA+CD27−CD4+T cells, low levels of ATP activate AMPK, inducing a constitutivep38 expression. Glucose deprivation induces AMPK and p38 activationin “non-senescent”CD27+T cells, leading to a decrease in both telo-merase activity and proliferation, mimicking the phenotype of highlydifferentiated T cells. Inhibition of p38 signaling promotes autophagyand restores energy-dependent functions in these cells (Henson et al.,2014). In summary, these results suggest a strong link between T cellaging and metabolic energy status.3. The impact of cytomegalovirus (CMV) on T cell agingCytomegalovirus (CMV) is a highly prevalent, persistent β-herpes-virus virus establishing lifelong latent infection within the host.Depending on socioeconomic factors and the geographic region, CMVinfects 40–100% of adult populations (Mocarski and Courcelle, 2001).By the age of 40, between 50 and 85% of adults are seropositive for thevirus (Selinsky et al., 2005). The primary infection and reactivations areusually asymptomatic in immunocompetent persons, but chronic CMV-infection contributes to age-related changes of the immune system andaccelerates some aspects of immunosenescence (Arnold et al., 2011;Brunner et al., 2011). CMV infection is mainly controlled by cytotoxicCD8+T cells, but CD4+T cells also play an important role in the an-tiviral response. In order to keep the virus under control, a high fractionof the adaptive immune compartment is required. Indeed, it is believedthat healthy middle-aged people use 10% or more of their circulatingCD4+and CD8+T cells to recognize CMV antigens and control viralproliferation (Sylwester et al., 2005).
This represents a consistent stress for the body, and leads to dysfunction in adaptive and innate immunity,thus contributing to immunosenescence. The frequency of CMV-specificT cells increases with each cycle of viral reactivation leading to anaccumulation of high numbers of CMV-specific, senescent-like CD8+Tcells, while the amount of circulating naïve T cells is reduced (Karreret al., 2003;Northfield et al., 2005) This process has been defined as“memory inflation“, and represents one of the features of im-munosenescence. As memory inflation dramatically affects the T cellphenotype by driving adaptive immune cells to terminal differentiation,an impact of CMV on immune responses and diseases should be ex-pected. In the elderly, high concentrations of CMV-specific antibodieshave been linked to increased mortality (Simanek et al., 2011;Wanget al., 2010). Furthermore, CMV seropositivity has been shown tocontribute to reduced survival in a cohort of Swedish octa- and non-agenarians (Wikby et al., 2005), and some studies showed a link be-tween CMV and a variety of chronic diseases with an inflammatorycomponent such as cardiovascular diseases (Du et al., 2018;Wang et al.,2017b). In CMV seropositive people, the increased production of pro-inflammatory cytokines in vascular smooth muscle cells may contributeto inflammation-mediated vascular pathology (Qiu et al., 2008). Anegative effect of CMV on the responses to pathogens and vaccines hasalso been reported in some studies. CMV seropositivity and antibodylevels are associated with lower antibody production after influenzavaccination in young and older persons (Frasca and Blomberg, 2016;Trzonkowski et al., 2003) Five years after booster vaccination againstdiphtheria, the concentrations of diphtheria-specific antibodies arelower in CMV seropositive compared to CMV seronegative individuals(Weinberger et al., 2017). In mouse models, aged MCMV-positive micehave limited T cell responses to influenza vaccines and infection com-pared to MCMV-negative mice (Mekker et al., 2012). However, severalstudies also reported the absence of a CMV-related effect on vaccine-induced immune responses (van den Berg et al., 2019). A recent reporthas shown that, in old mice infected with MCMV early in life, thepresence of the virus may have positive impacts on adaptive hetero-logous immunity (Smithey et al., 2018). Thus, latent persistent CMVmay modulate adaptive immunity in a manner that, in some conditions,may benefit host defense. Overall, the massive impact of CMV on thecomposition of the T cell compartment is undisputed, but the evidencefor detrimental effects on immune responses to other antigens and onoverall health or mortality is more controversial and needs further in-vestigation (Jergovićet al., 2019).
4. T cells, aging and senescence: CD28−T cells
Particularly within the CD8+T cell compartment, repeated ex-posure to antigens leads to the accumulation of highly differentiated Tcells. CD8+T cells play an important role in the recognition andclearance of cells infected by intracellular pathogens. Upon TCR re-cognition of MHC-I-bound peptides presented by antigen presentingcells (APC), CD8+T cells receive an activation signal. For full activa-tion, T cells need a second signal known as costimulatory signal. One ofthe strongest and best-defined co-stimulatory signals is provided by theinteraction of CD28, expressed on the surface of T cells, and its ligandsCD80 or CD86 on the surface of activated APC. Fully activated naïve Tcells start to proliferate and concomitantly differentiate into effector Tcells, which die by apoptosis after performing their effector functions,and memory T cells, which are retained for recall responses against thesame antigens in case of a subsequent exposure. Under persistent an-tigenic stimulation, several activation cycles occur. With repetitiverounds of stimulation and proliferation, CD28 expression is progres-sively downregulated on the surface of CD8+T cells, leading to theaccumulation of CD28−T cells. Although both CD8+and CD4+T cellsare affected, the downregulation of CD28 is more evident in CD8+Tcells (Qin et al., 2016). Loss of CD28 expression is associated with therapid inactivation of telomerase, and therefore CD8+CD28−T cellsexhibit shorter telomeres in comparison to CD8+CD28+T cells(Monteiro et al., 1996). In addition, CD28−T cells show reduced an-tigen-induced proliferation and enhanced production of pro-in-flammatory molecules (Fagnoni et al., 1996;Posnett et al., 1994).Nevertheless, it has been demonstrated that CD28−T cells can stillproliferate under certain conditions (Vivar et al., 2011;Weng et al.,2009). In particular, CD8+CD28−T cells are able to proliferate whenco-stimulation is provided by cytokines, such as IL-15 (Brunner et al.,2012;Chiu et al., 2006). As one of the main hallmarks of cellular se-nescence is a permanent and irreversible cell cycle arrest, CD28−T cellscannot be considered truly senescent cells. Despite this, it is correct tosay that this subset represents a population of highly differentiatedcells, with a phenotype somewhat similar to “senescent-like”cells.Accumulating evidence indicates that CD8+CD28−T cells are asso-ciated with numerous disorders, regardless of age. Increased numbers ofCD8+CD28−T cells can be found in tumor microenvironments and thePB of cancer patients, and this elevation is associated with malignancy(Tsukishiro et al., 2003). Accumulation of CD8+CD28−T cells is alsoobserved in chronic viral infections, such as CMV, in both PB and BM(Derhovanessian et al., 2011;Pangrazzi et al., 2017b). In the elderly, increased frequencies of CD8+CD28−T cells can be observed in theperiphery (Fagnoni et al., 1996). The accumulation of this subset ofhighly differentiated CD8+T cells has been associated with a reducedimmune response to pathogens and vaccines in old age (Saurwein-Teisslet al., 2002). Additionally, the frequency of this population was shownto increase in the BM during aging (Naismith et al., 2019;Naismith andPangrazzi, 2019;Pangrazzi et al., 2017a). It has been reported thatCD8+CD28−T cells are detrimental for elderly people, as their in-creased frequency is associated with high inflammation levels and riskof age-related diseases and mortality (Wikby et al., 2005). Therefore,due to their pro-inflammatory phenotype, CD8+CD28−T cells maycontribute to chronic low-grade inflammation observed in old age (in-flammaging).(Franceschi et al., 2018;Naismith et al., 2019;Pangrazziet al., 2017a). It can be concluded that, although CD28−T cells may notbe fully senescent, indications exist that the presence of this populationis detrimental and may contribute to diseases. CD28−T cells are veryheterogenous and therefore it is important to define specific markers toidentify a “truly senescent”population within CD28−T cells.
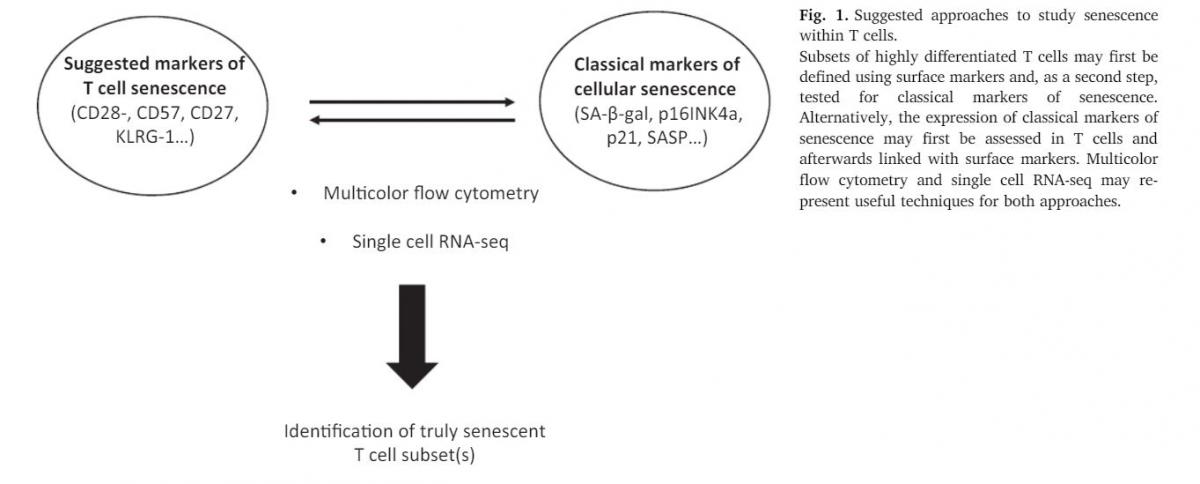
5. T cells, aging and senescence: searching for the “truly”senescent T cell population
During their differentiation process, both CD8+and CD4+T cellsgain the expression of CD57 (Brenchley et al., 2003;Unemori et al.,2009). It has been suggested that T cells may first lose CD28 and, as asecond step, express CD57, therefore generating a population ofCD28−CD57+T cells which is more terminally differentiated (Striogaet al., 2011). The expression of pro-inflammatory molecules, such asIFNγand TNF, and levels of ROS, commonly associated with senes-cence, are higher in CD28−CD57+CD8+T cells compared toCD28+CD57−and CD28−CD57−CD8+T cells. In contrast, gene ex-pression related to DNA repair and apoptosis, as well as the prolifera-tion rate, are reduced in comparison to the other subsets. These resultsindicate that CD28−CD57+CD8+T cells are at least one step closer tosenescent cells than CD28−T cells (Pangrazzi et al., 2019).
There are indications that the expression of CD27 may allow theidentification of CD8+CD28−T cells that are closest to terminal dif-ferentiation and senescence (Plunkett et al., 2007). Indeed, it has beenreported that CD27−CD28−CD8+T cells have shorter telomeres thanCD27+CD28−and CD27−CD28+CD8+T cells, with decreased telo-merase activity and reduced proliferation. This suggests thatCD28−CD27−CD8+T cells had differentiated to a point, in which co-stimulatory signals are no longer sufficient to induce telomerase ac-tivity, like it is happening for CD27+CD28−and CD27−CD28+CD8+T cells (Arnold et al., 2011). Thus, a differentiation pathway of CD8+Tcells from early differentiated CD28+CD27+to differentiatedCD28−CD27+and to terminally differentiated CD28−CD27−T cellswas suggested. Similar to CD28−T cells, CD27−CD28−T lymphocytesaccumulate in the elderly and in the presence of chronic viral infections(Fletcher et al., 2005). As CD8+T cells may first lose CD28 and, as asecond step, CD27, this model can explain why CD8+CD28−T cellsmay still be functional. In contrast to CD8+T cells, CD27 seems to bedown-regulated before CD28 in CD4+T cells (Kovaiou et al., 2005;Moro-García et al., 2013).
Another approach has been tested, in order to identify the “most”senescent T cell candidate, in this case within CD4+T cells (Hensonet al., 2014). It was observed that CD45RA+CD27−CD4+T cells dis-played mitochondrial dysfunction, another typical feature of senes-cence, which could be reversed after blocking p38 mitogen-activatedprotein kinases (MAPK).
Senescent cells are known to secrete a broad spectrum of moleculessuch as cytokines, proteases and growth factors, which distinguish themfrom healthy cells present within tissues. These factors represent a ty-pical hallmark of senescence and therefore they have been defined assenescence-associated secretory phenotype (SASP) (Ghosh and Capell,2016;Lopes-Paciencia et al., 2019). It has been reported thatCD45RA+CD27−CD4+T cells may show the typical SASP phenotype,with high secretion of pro-inflammatory cytokines, chemokines, growthfactors and proteases (Callender et al., 2018). Interestingly, MAPK ac-tivity was necessary to induce the SASP, as it could partially be revertedby inhibiting p38 MAPK activity. However, it must be taken into con-sideration that CD45RA+CD27−T cells represent a very heterogeneouspopulation, which includes CD28+/−and CD57+/−cells. Very re-cently, our group showed that a SASP phenotype may also be presentalso in peripheral CD28−CD57+CD8+T cells (Pangrazzi et al., 2019).Despite this, no T cell population has been identified, which shows allhallmarks of senescence, including for example upregulation of the pro-inflammatory molecules IL-6 and IL1βand all proteases and growthfactors associated with senescence in fibroblasts. Thus, in order tobetter define senescent T cell candidates, and to understand whethersenescent T cells are truly senescent according to the classical defini-tion, a combination of several markers is essential. Most likely thesecells can be found within CD28−CD57+CD45RA+CD27−populations,but additional markers need to be identified, and they might be dif-ferent for CD4+and CD8+T cells.
6. The interplay between T cells and other senescent cells
The accumulation of senescent cells has been shown to contribute toage-related diseases and aging itself (Baker et al., 2016;Xu et al., 2018).Induction of the DNA damage response (DDR) is known to support theactivation of the innate and adaptive immune system (Nakad andSchumacher, 2016;Xu, 2006) Usually, activation of the DDR also in-itiates a senescence program (Burton and Faragher, 2015;D'Adda DiFagagna, 2008). During the initiation of the cellular senescence pro-gram, the DDR is important for inducing the SASP, which is capable ofattracting and activating innate and adaptive immune cells via cyto-kines and chemokines. These molecules can support innate immuneresponses through several pathways, including the upregulation ofNKG2D (natural killer group 2, member D) ligands and interferon-regulatory factors. Thus, NK cells, natural killer T (NKT) cells, γδ Tcells, dendritic cells and monocytes, but also T and B cell subsets, can beactivated by products of the SASP. We can speculate that, as at leastsome SASP molecules are expressed by subsets of highly differentiatedT cells (Callender et al., 2018;Pangrazzi et al., 2019), the presence ofsenescent T cells may influence neighboring cells, such as immune cellsor other cells within tissues.Recently, it has been reported that senescent dermal fibroblastsexpress HLA-E. Expression of HLA-E is also higher on senescent cellsfrom old compared to young donors. HLA-E binds the inhibitory re-ceptor NKG2A expressed by highly differentiated CD8+T cells and NKcells (Pereira et al., 2019). The acquisition of NK receptors by highlydifferentiated T cells may be due to epigenetic mechanisms such asmethylation of certain genes, as reported for NK cells (Wiencke et al.,2016). NKG2A levels are higher on CD27−CD28−, compared toCD27+CD28−and CD27+CD28+CD8+T cells. Thus, when highlydifferentiated T cells are present, NKG2A-HLA-E binding occurs, andprevents the elimination of senescent cells by cytotoxic T cells. Inter-estingly, when the interaction between HLA-E and NKG2A is blocked,CD8+T cell mediated cytotoxicity against senescent cells is boosted(Pereira et al., 2019).In summary, recent studies suggest that senescent-like T cells play amajor role in the accumulation of senescent cells within tissues.Although it is still not clear whether some T cell subsets may be se-nescent, the clearance of highly differentiated T cells may represent apromising strategy to fight cellular senescence in other cell types, suchas fibroblasts and pre-adipocytes.
T O A C C E S S T H E R E S T O F T H E S T U D Y, P L E A S E V I S I T T H E S O U R C E .
.















