Hi AgeVivo, Caliban,
Thanks for pointing me to this post – I had not seen it.
No, I do not think anything has changed much since that post in 2008 and I do agree with much of what Mr. Rae has said – although not with all of it (but when do scientists ever agree totally, see below).
Some points regarding the assessment of the 2008 paper by da Silva et al.:
1) I am not sure that there is strong evidence that the mice used in the paper are unreasonably and worryingly short lived.
The citation given in support of the notion that this is the case clearly illustrates that different mouse strains have very different expected longevities.
It does not, however, give data specifically on the female Swiss Webster outbred albino mice used by da Silva et al. So the question is, what is the “normal” mean and maximum lifespan of this specific type of WT mice.
I do not work in mouse lifespan studies myself and I do not know the normal lifespan for the specific strain used in the paper (if anybody has hard data on this issue, it would be useful indeed). However, the paper by da Silva cites one other publication [1], which seems to support their assertion that the lifespan observed is pretty normal for females of this type of outbred mice.
Furthermore, the longest surviving mice in the study lived to about 140 weeks (980 days or almost 2.7 years). Compared to other published studies on mouse lifespan, 140 weeks does not strike me as suspiciously short lived – unless somebody has a reference regarding this particular type of mice that says otherwise.
The method section of the da Silva paper also indicates careful animal care protocols including specific pathogen free conditions.
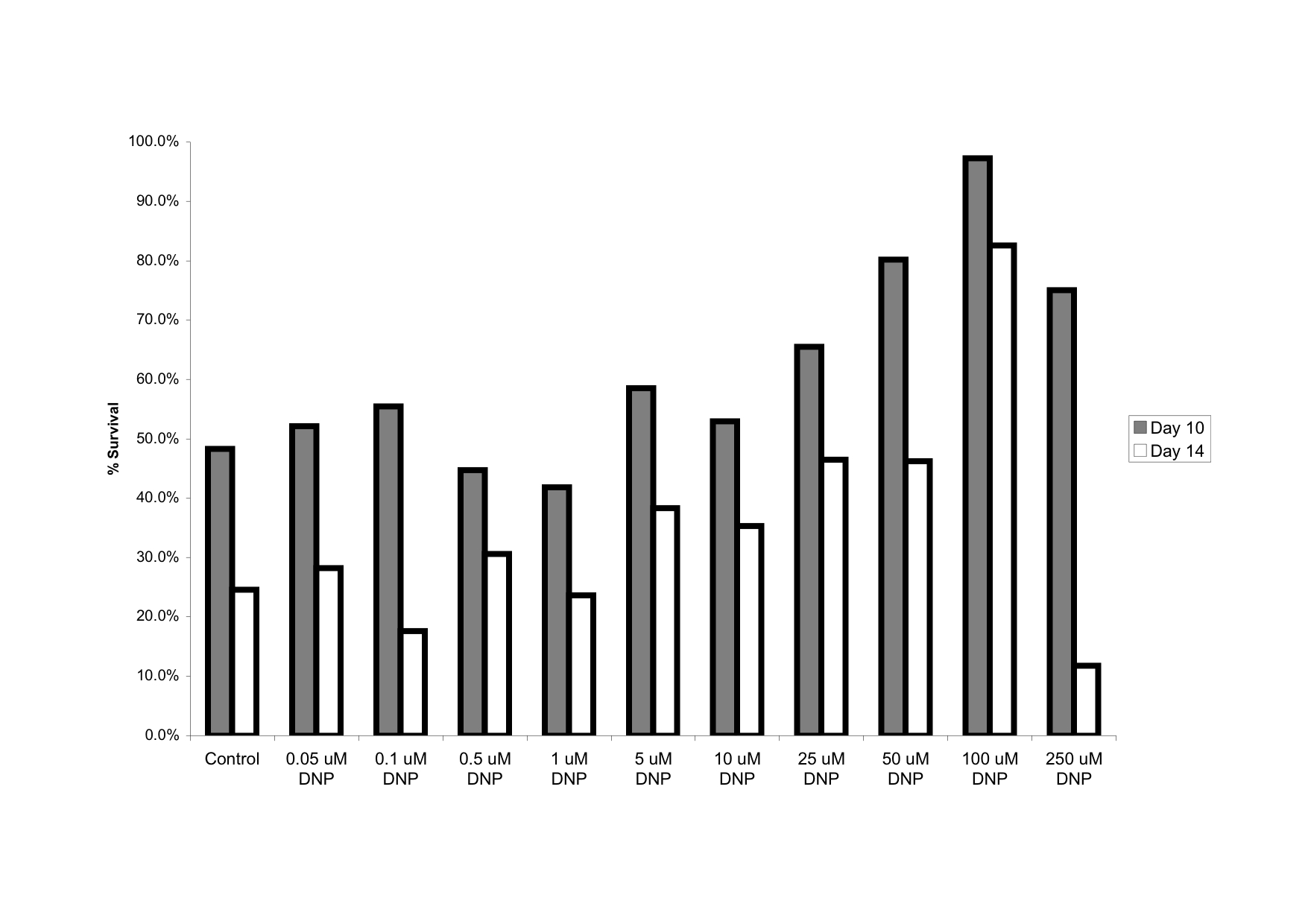
2) Maximum lifespan is commonly measured as the average of the longest surviving 10% of a cohort. By this measure, there clearly is at last a trend to increased maxLSP with DNP treatment (Figure 4 in da Silva et al.). Whether this is statistically significant is not clear but this is probably at least partially a question of n-numbers (30 animals in each group – so the top 10% are only 3 individuals). Mouse lifespan studies are expensive and n-numbers are often regrettably small (one of the reasons to consider working with simple model organisms, IMHO).
I guess my point here is that the judgment that this work is in some way “nonsense” is maybe not well supported by evidence in the original post. In fact, I think it is quite a good paper on this approach, although I agree that they may be overstating the CR case a little in the discussion.
3) I also fully agree that there is little clear evidence that DNP has slowed ageing or mimicked CR in these animals. Showing effect on ageing would require not just clear extension of maximum lifespan but ideally evidence for a higher mortality doubling times (lower ageing rate). Assessing mortality patterns is very difficult (expensive) to do in mice because of the large number of animals that would be required. This is, however, one thing that is fairly easy to do in nematodes where 100s or even 1000s of animals can be used relatively easily (and cheaply).
4) I do further agree that there are many open questions regarding “what happened”, in particular regarding the mechanism of action of DNP in these mice. A reduction in body fat certainly is one possibility here. Also, it is interesting to note that DNP affects the brain in particular and that it is here that the markers of oxidative damage are most changed. This is interesting but also represents a challenge, as different organs are affected differently.
5) Most of all, I emphatically agree that DNP is a dangerous drug and should not be viewed as lifespan extension medication for humans.
Does this mean that there is no reason to carry out the work we have proposed ?
Clearly, I do not think so - our proposal is very explicitly a basic science proposal and we consider DNP an interesting proof-of-principle compound (laboratory reagent) for testing a specific “metabolic tuning” approach in the context of the mitochondrial free radical theory of ageing (mFRTA). This work is also intended to test aspects of the mFRTA itself (which has been much debated and challenged recently).
The fact that DNP has already been shown to extend lifespan in mice just makes it even more interesting to figure out how it might (or might not) work.
Unlike da Silva et al. we have the luxury of doing in vivo ROS dose-response curves first, in order to get an ideal dose to work with. We can also carry out studies big enough to evaluate maximum lifespan and maybe even mortality changes. We routinely observe parameters of mitochondrial function and damage and we will have to worry less about complex pharmacokinetics effect.
If we can use DNP to significantly reduce oxidative damage to mitochondria (without other obvious detrimental changes) and this does NOT lead to maximum lifespan extension / slower ageing rate – that, in my view, would be a serious challenge to the free radical theory of ageing. If on the other hand chemical uncoupling can be used to reduce damage and preserve function (in this simple model organism), it would lend support to this avenues of research and might ultimately lead to compounds much safer than DNP. Again, the fact that there is already evidence that this approach has at least some beneficial effects, even in higher organisms, is actually very encouraging.
To summarize, I think the paper by da Silva et al. has illustrated that mild chemical uncoupling might be an interesting strategy for modulating lifespan. I do not think that there are problems with their work so severe to suggest that there is nothing further to explore. However, I fully agree that many important questions remain unanswered, which is why I wrote the proposal in the first place 8)
I hope this goes some way towards answering both AgeVivo’s and Caliban’s concerns / comments – let me know if it does not ;-)
[1] Guayerbas N, De La Fuente M (2003) An impairment of phagocytic function is linked to a shorter life span in two strains of prematurely aging mice. Dev. Comp. Immunol. 27, 339–350.
Edited by Jan Gruber, 17 August 2010 - 05:17 PM.

































 ,
, 
 JanGruber.doc 20KB
149 downloads
JanGruber.doc 20KB
149 downloads