































.
F U L L T E X T S O U R C E : nature
Abstract
Nutrients are absorbed solely by the intestinal villi. Aging of this organ causes malabsorption and associated illnesses, yet its aging mechanisms remain unclear. Here, we show that aging-caused intestinal villus structural and functional decline is regulated by mTORC1, a sensor of nutrients and growth factors, which is highly activated in intestinal stem and progenitor cells in geriatric mice. These aging phenotypes are recapitulated in intestinal stem cell-specific Tsc1 knockout mice. Mechanistically, mTORC1 activation increases protein synthesis of MKK6 and augments activation of the p38 MAPK-p53 pathway, leading to decreases in the number and activity of intestinal stem cells as well as villus size and density. Targeting p38 MAPK or p53 prevents or rescues ISC and villus aging and nutrient absorption defects. These findings reveal that mTORC1 drives aging by augmenting a prominent stress response pathway in gut stem cells and identify p38 MAPK as an anti-aging target downstream of mTORC1.
Introduction
The incidence of aging-related disorders is increasing due to aging of the population1. Nutrient malabsorption is common among the elderly, and often causes anemia and other illnesses2. Nutrients are absorbed by the intestinal villi, which are composed of a layer of intestinal epithelial cells (IECs) and the lamina propria, and the absorption activity is affected by the size and density of villi3. The epithelial layer is renewed every 4–5 days by Lgr5+ intestinal stem cells (ISCs) following a symmetric division and neutral shift pattern4,5,6, which generate transient amplifying (TA) progenitor cells that later differentiate into absorptive or secretory cells7. The densities of crypts and villi are controlled by ISC fission8. In addition, Bmi1+ quiescent stem cells, TA cells, and secretory progenitors can revert to Lgr5+ ISCs after damage to regenerate the villi9,10,11,12,13. It has been reported that the number of ISCs and ISC regenerative activities are decreased in 17–24-month-old mice14,15, yet whether aging affects villus function and how villus aging is controlled remain less well understood.
mTOR, a sensor of nutrients and growth factors, is a central regulator of aging and a target for lifespan and healthspan extension16,17,18,19,20. mTOR forms mTORC1 and mTORC2 complexes, and mTORC1 activation promotes cell proliferation by increasing global protein synthesis and other anabolic processes21,22,23,24, which is assumed to cause irreversible cell senescence and/or oxidative and proteostatic stress and thus aging25,26,27,28. However, this hypothesis is not consistent with the reversible aging theory29,30. mTORC1 signaling has been shown to be required for IEC proliferation during homeostasis and regeneration31,32,33,34, including regeneration mediated by quiescent ISCs35,36. In addition, several studies have shown that diet restriction promotes Lgr5+ ISC expansion via mTORC1 signaling, although conflicting results have been reported regarding the exact roles played by mTORC114,37,38. Nonetheless, the mechanisms by which mTORC1 signaling regulates ISCs and villus aging warrant further investigation.
The current genetic study reveals that mTORC1, which is hyperactivated in IECs, especially ISCs and TA cells of aged mice, drives villus aging by inhibiting ISC/progenitor cell proliferation through amplifying the MKK6-p38-p53 stress response pathway, with MKK6 protein synthesis being upregulated by mTORC1 activation. Villus aging can be slowed or prevented by targeting p38 MAPK or p53. These genetic results establish a prominent stress response pathway as an mTORC1 downstream effector in aging control, elucidate the mechanisms governing villus aging, and identify p38 MAPK as an antiaging target downstream of mTORC1 signaling.
Results
Aging causes a decrease in the numbers of crypts and TA cells
To understand aging of the nutrient-absorbing organ, we collected small intestines from normal B6 mice of various ages, which were bred and housed in the same facility. Histological analysis revealed that 16 or 17.5-month-old mice, compared with 2-, 3.5-, 8-, and 12-month-old mice, showed significant decreases in the height and number of villi in the proximal and distal jejunum (Fig. 1a; Supplementary Fig. 1a, b). The 17.5-month-old mice showed a 12% increase in body weight compared with 2–3-month-old mice. Mice at 24 months of age also showed villus structural deterioration (Supplementary Fig. 1c). These results indicate a decline in the absorption surface in aged mice, which are disagreeable with a recent study by Nalaparenddy et al. reporting an increase in the villus size in old mice15. While the causes of this discrepancy can be complex, one explanation could be that Nalaparenddy’s study used young and old mouse cohorts purchased from different suppliers15. We found that 16- or 17.5-month-old mice showed decreases in nutrient-absorbing activities for unmetabolizable L-glucose, amino acids, and fatty acids (Fig. 1b; Supplementary Fig. 1d).
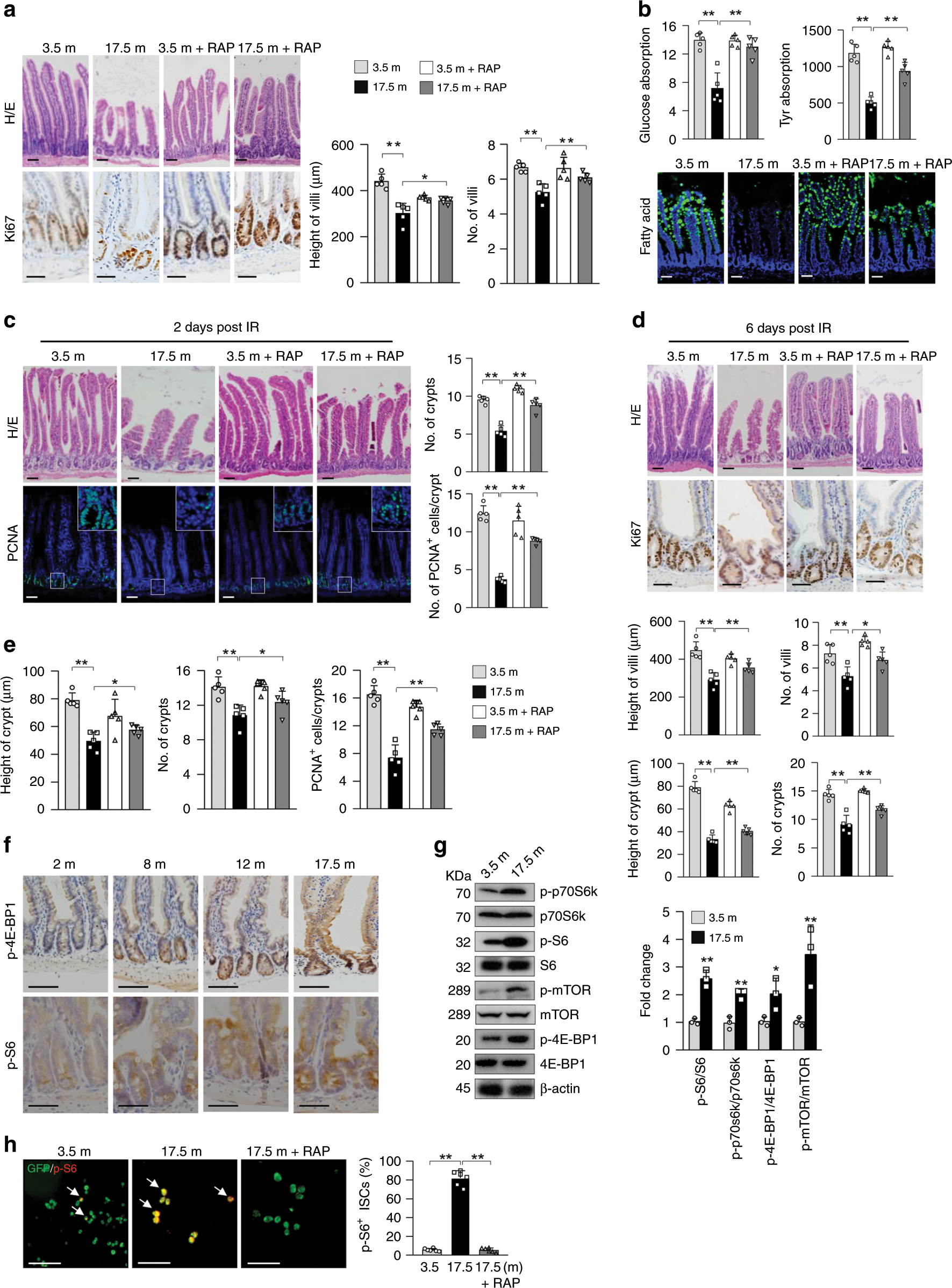
Fig. 1: Enhanced mTORC1 activation in ISCs/TA cells of geriatric mice drives villus natural aging.
a–d Seventeen and half-month-old mice showed deterioration in villus and crypt structures (right panels: quantitation data) (a), decreased nutrient absorption activities (b), increased sensitivity to IR-induced decreases in the numbers of crypts and proliferating cells at day 2 post IR ©, and compromised regeneration (decreases in the height and number of villi and crypts) at day 6 (d) compared with young mice, which were partially rescued by 1.5 months of RAP treatment (3 mg/kg body weight) starting at 16 months of age. Data are expressed as mean ± SEM. N = 5 mice per group. *P < 0.05, **P < 0.01 (determined using Student’s t test). e Seventeen and half-month-old mice showed decreases in the height and number of crypts and the number of proliferating TA cells (based on (a)), which were rescued by RAP. Data are expressed as mean ± SEM. N = 5 mice per group. *P < 0.05, **P < 0.01 (determined using Student’s t test). f Representative images (proximal jejunum midline sections) showed that mTORC1 activation was increased with age in crypt cells. g Western blot results showed that mTORC1 activation was increased in the crypt samples of 17.5-month-old mice compared with 3.5-month-old mice. Isolated crypts were directly lysed and used for WB analysis. Data are expressed as mean ± SEM. N = 3 mice per group. *P < 0.05, **P < 0.01 (determined using Student’s t test). h More Lgr5+ ISCs isolated from 17.5-month-old mice showed mTORC1 activation than those from 3.5-month-old mice, which was suppressed by RAP treatment. Lgr5+ ISCs were isolated from the small intestines of Lgr5-GFP-CreERT mice with FACS sorting and stained for p-S6. Right panel: quantification data (mean ± SEM). N = 6 mice per group. **P < 0.01 (determined using Student’s t test).
The old mice also showed increased sensitivity to ionizing radiation (IR), manifested by greater decreases in the numbers of crypts and proliferating cells and greater increase in apoptotic cells than young mice at day 2 post IR (Fig. 1c; Supplementary Fig. 2a). This was associated with a decrease in PCNA and cyclin E and an increase in p53 in crypt samples (Supplementary Fig. 2b). Similar results were obtained at day 3 post IR (Supplementary Fig. 2c). The increased damage in old mice may be the cause of compromised villus regeneration observed at day 6 post IR, manifested by decreases in the height and number of villi and crypts (Fig. 1d; Supplementary Fig. 2d). Overall, these findings indicate that aging is associated with a deterioration of villus structure and function, increased sensitivity to stress, and compromised regeneration.
Old mice also showed decreases in the height and number of crypts, the ISC/progenitor-containing glands that control villus size and density. We observed a decrease in the number of Ki67+ progenitor cells (Fig. 1a, e; Supplementary Fig. 1a), but no significant changes in the numbers of apoptotic or senescent cells or differentiation of villus cells after normalized to the villus size (Supplementary Fig. 2e–g). Although the numbers of villi and crypts were decreased in old mice, the crypt-to-villus ratio was unaltered (Supplementary Fig. 2h), suggesting that aging-related decreases in villus height and density may be caused by decreases in the numbers of proliferating TA cells and crypts, respectively. Yilmaz’ group also reported that aging caused decreases in the number and regeneration ability of ISCs, but not defect in goblet or enterocyte differentiation14. Taken together, our and others’ studies suggest that villus aging-associated decrease in villus size and density are likely caused by defects in ISCs and TA progenitors14,15.
Hyperactivated mTOR in IECs contributes to villus aging
mTOR activation is implicated in the aging process34,39. Immunostaining showed that p-4E-BP1 and p-S6, indicators of mTORC1 activation, were increased with age in IECs, especially in crypt cells (Fig. 1f). Western blot analysis confirmed an increase in mTORC1 activation in crypt samples of old mice (Fig. 1g). Because the number of ISCs was too limited for western blot analysis, we sorted Lgr5+ ISCs from 3.5- and 17.5-month-old Lgr5-GFP-CreERT mice and immunostained them for p-S6. We found that substantially more ISCs displayed mTORC1 activation in old mice (Fig. 1h). The factors that cause mTORC1 hyperactivation in aged IECs and ISCs may include systemic and niche cues and, warrant further investigation39,40.
Interestingly, treatment of 16-month-old mice with rapamycin (RAP), an mTORC1 inhibitor21, for 1.5 months inhibited mTORC1 activity (Supplementary Fig. 2i) and partially rescued aging-like phenotypes, including decreases in villus height and density and nutrient absorption activities, increased sensitivity to IR at day 2, and compromised villus regeneration at day 6 post IR (Fig. 1a–d; Supplementary Fig. 2a–d). It has been previously reported that acute RAP inhibited villus regeneration in normal mice31,33,36. Here, we carried out the above experiments 3 days after completion of drug treatment to avoid possible acute effects of RAP. Moreover, RAP mainly rescued IR-induced decrease in cell proliferation and increase in apoptosis at day 2 or 3 post IR (Fig. 1c; Supplementary Fig. 2a–c), which might help villus regeneration. In addition, RAP partially rescued the decreases in the height and number of crypts and the number of proliferating TA cells in old mice (Fig. 1a, e). These results suggest that enhanced mTORC1 activation contributes to villus natural aging.
Tsc1 ablation in IECs causes villus premature aging
To test whether mTORC1 activation causes villus aging, we ablated Tsc1, a disease gene encoding an mTOR suppressor, using Villin-Cre mice. Villin is expressed in all small intestinal IECs, including ISCs, but not colon crypts41. Villin-Cre;Tsc1f/f mice exhibited enhanced mTOR activation in the whole villi (Supplementary Fig. 3a). We found that 2- or 3-month-old Villin-Cre;Tsc1f/f mice showed increases in the number and size of crypts, but insignificant increases in villus size (Supplementary Fig. 3b). The nutrient absorption rates were not altered in these mice (Supplementary Fig. 3c), suggesting that mTORC1 activation in IECs per se does not affect nutrient absorption in the presence of normal villi. Surprisingly, 4–6-month-old mutant mice showed villus/crypts similar to age-matched wild-type mice (Supplementary Fig. 3b).
Starting from 7 months of age, the mutant mice displayed aging-related changes, including decreases in villus height and density, nutrient absorption activities, and villus regeneration (Supplementary Fig. 3b–d). The compromised regeneration in Villin-Cre;Tsc1f/f mice was likely caused by increases in IR-induced cell death and villus damage at day 2 post IR (Supplementary Fig. 3d). Overall, these results indicate that Tsc1 deletion in IECs cause villus premature aging.
Tsc1 ablation in ISCs-induced villus aging is rescued by RAP
The above studies show that nutrient absorption defects observed in villus natural or premature aging models are associated with decreases in villus size and density, whereas mTORC1 activation per se in IECs does not affect nutrient absorption activities at young age (Supplementary Fig. 3c). Since villus size and density are controlled by ISC and progenitor cells, we ablated Tsc1 in ISCs using Lgr5-CreERT mice4. Lgr5-CreERT reportedly only labels part of ISCs after tamoxifen (TAM) administration and generates chimeric labeling patterns4. Our lineage-tracing studies revealed that three doses of TAM administered daily to 1-month-old Lgr5-CreERT;tdTomato mice labeled 65% of the crypts by 2 months of age (Supplementary Fig. 4a). A majority of the cells in almost all the villi were Tomato-labeled, likely because each villus contains at least six crypts7.
.../...
F O R T H E R E S T O F T H E S T U D Y, P L E A S E V I S I T T H E S O U R C E .
.
















