































.
O P E N A C C E S S S O U R C E : nature
Abstract
Aging is characterized by the loss of homeostasis and the general decline of physiological functions, accompanied by various degenerative diseases and increased rates of mortality. Aging targeting small molecule screens have been performed many times, however, few have focused on endogenous metabolic intermediates—metabolites. Here, using C. elegans lifespan assays, we conducted a worm metabolite screen and identified an eukaryotes conserved metabolite, myo-inositol (MI), to extend lifespan, increase mobility and reduce fat content. Genetic analysis of enzymes in MI metabolic pathway suggest that MI alleviates aging through its derivative PI(4,5)P2. MI and PI(4,5)P2 are precursors of PI(3,4,5)P3, which is negatively related to longevity. The longevity effect of MI is dependent on the tumor suppressor gene, daf-18 (homologous to mouse Pten), independent of its classical pathway downstream genes, akt or daf-16. Furthermore, we found MI effects on aging and lifespan act through mitophagy regulator PTEN induced kinase-1 (pink-1) and mitophagy. MI’s anti-aging effect is also conserved in mouse, indicating a conserved mechanism in mammals.
Introduction
Molecules that promote healthy aging have been searched and studied because of their potential to target human aging1. The aging process is closely linked to metabolism2. Metabolism is composed of chemical reaction networks, which can be represented with metabolites as nodes and enzymes as edges3. Metabolites, such as NAD+ and related metabolites4,5,6, spermidine7, pyruvate8, N-acetylglucosamine (GlcNAc)9, and α-ketoglutarate10, have been reported to alleviate aging in model organisms.
The number of metabolites varies in magnitudes across the four kingdoms: tens of thousands in plants, thousands in humans, and one thousand in worm11,12,13. To investigate the influence of available metabolites on C. elegans lifespan, we conducted a preliminary screen on worm 100 metabolites and found that an evolutionarily conserved metabolite, myo-inositol (MI), significantly increased lifespan across a large range of concentrations.
MI is an extensively studied metabolite14,15,16,17, whose isomers or derivatives participate in many fundamental physiological processes18. Among its nine possible stereo-isomers, MI is the most abundant18. MI is a common ingredient in food19, and it is also used as a food supplement to alleviate a variety of diseases20, including diabetes21 and polycystic ovary syndrome (PCOS)22. An isomer of MI, D-chiro-Inositol, which does not exist endogenously in either C. elegans or Drosophila melanogaster, has been reported to extend fly lifespan through unknown mechanisms and was speculated to act through dFOXO13, 23. Inositol in the form of MI itself has not been reported to influence aging.
There are scores of MI derivatives, classified into three groups, phosphatidylinositol phosphates (PIs), inositol phosphates (IPs), and glycosyl phosphatidylinositols (GPI)24,25,26. Among them, abnormally high levels of PI(3,4,5)P3, a key lipid component in the insulin and insulin-like growth factor signaling pathway (IIS), is known to promote cancer and shorten lifespan through activation of AKT in mouse/mammals27. Conversely, another derivative, IP7 is reported to inhibit AKT in mouse28, but has not been reported to influence aging. Both PI(3,4,5)P3 and IP7 in aging were indirectly studied by perturbations of their synthases, i.e., loss of function (LOF) of worm Phosphatidylinositol 3-kinase (age-1)29, mouse liver phosphatidylinositol-4,5-bisphosphate 3-kinase (Pi3k)30 and LOF of inositol hexaphosphate kinase (Ip6k1) in mouse28, respectively. It remains unknown whether or how the other derivatives influence aging.
Here we found that MI extended worm lifespan and promoted healthspan, in other words, prevented the age-related decline in physiological functions. Genetic dissection of the MI metabolic pathway suggests that MI alleviates aging through its derivative PI(4,5)P2, despite MI and PI(4,5)P2 being precursors of PI(3,4,5)P3. Moreover, its effects are completely dependent on the well-known tumor suppressor gene, daf-18 (phosphatase and tensin, Pten homologue), which has many functions besides being an enzyme in the MI metabolic pathway, and partially depends on its downstream mitophagy regulator pink-1 rather than its classical downstream daf-16. The anti-aging effects of MI were recapitulated in mice at phenotypic and transcriptome levels, suggesting a conserved mechanism in mammals.
Results
MI alleviates aging in worm
MI is a cyclohexanehexol (Fig. 1a) that is metabolized in some bacteria, most archaea, and all eukaryotes18. The major reactions in the MI pathway are conserved from worm, mouse to human. Here we define MI metabolic pathway as the reactions forming an unbroken chain with MI upstream or downstream as annotated by the three KEGG pathways: Inositol phosphate metabolism, Phosphatidylinositol signaling system and Glycosylphosphatidylinositol (GPI)-anchor biosynthesis.
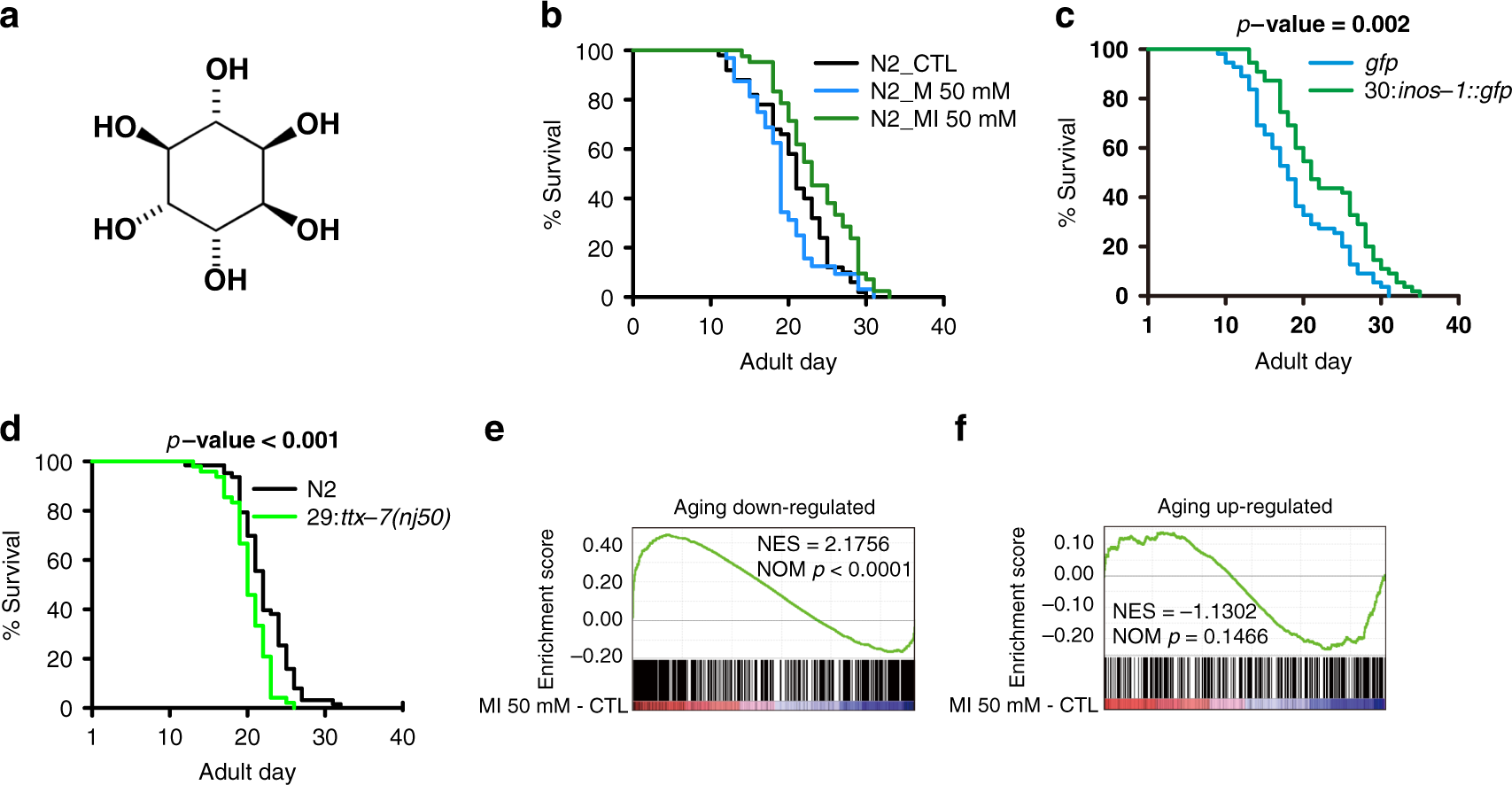
Fig. 1: MI promotes worm longevity.
a Chemical structure of MI. b Survival curves of N2 worms treated with or without 50 mM M (log-rank test p < 0.001) or MI (p < 0.01), N = 10. c Survival curves of inos-1 OE and control N2 worms (log-rank test p = 0.002). The number before “:” in a label indicates the reaction step in Fig. 4a, N = 11. d Survival curves of ttx-7 LOF and control N2 worms (log rank test p = 2.00E-04), N = 5. e, f GSEA results show worm aging downregulated or upregulated gene expressions measured by RNA-seq FPKM log2-fold-change, were reversely regulated by 50 mM MI (GSEA p < 0.0001 for aging downregulated genes and p = 0.1466 for aging upregulated genes, respectively), N = 1. Source data are provided as a Source Data file and Supplementary Dataset 1.
Through a small molecule screen with endogenous metabolites, we found that various concentrations of MI, from 1 up to 700 mM, and optimally at 500 mM, could extend worm lifespan (Supplementary Fig. 2d). To avoid the strong osmotic stress associated with these concentrations, we performed further analysis to delineate its molecular pathways at 50 mM MI in downstream experiments, which extended worm mean lifespan by 14% (p < 0.01, Fig. 1b). In addition, as osmotic effects, which can be induced by chemicals that alter osmolyte balance, like sorbitol and are found to extend worm lifespan31, we used 50 mM mannitol (M) as an osmotic control, because it cannot be further metabolized in worms.
To further validate the effect of MI on lifespan, we constructed a worm strain, over-expressing inositol 3 phosphate synthase (inos-1), the rate-limiting enzyme to synthesize MI in most species, including worms and mammals32 as a fusion protein with GFP, and backcrossed to N2 six times. We found that this inos-1::GFP overexpression (OE) strain had a 19% increase in lifespan compared with GFP only OE strain (p = 0.002, Fig. 1c).
The second step in the MI synthesis pathway is catalyzed by the enzymes inositol monophosphatase (TTX-7) or Y6B3B.5, to produce MI from the three types of inositol monophosphate (IP). IP is produced by INOS-1 or the degradation product of inositol phosphates. The ttx-7 LOF allele nj51, shortened worm lifespan by 8% (p < 0.001, Fig. 1d).
We further examined MI’s effect on the transcriptome changes. As expected, MI induced a transcriptome change opposite to the changes induced by aging, in particular, aging downregulated genes were significantly upregulated by MI (p < 0.0001, Fig. 1e), and aging upregulated genes showed a trend towards downregulation by MI (p = 0.1466, Fig. 1f).
MI promotes worm healthspan
To examine whether MI also promotes healthspan, we first scored worm mobility by counting body-bending rate and pumping rate in adult day (AD) 4 to day 10. Both MI treatment and inos-1 OE increased worm mobility (*p < 0.05 on AD_10 for MI, both AD_6 and AD_10 for inos-1 OE, Fig. 2a, b), and alleviated worm pumping-rate declines during aging (*p < 0.05 on AD_6 for MI or AD_8 for inos-1 OE, Supplementary Fig. 2a, b).
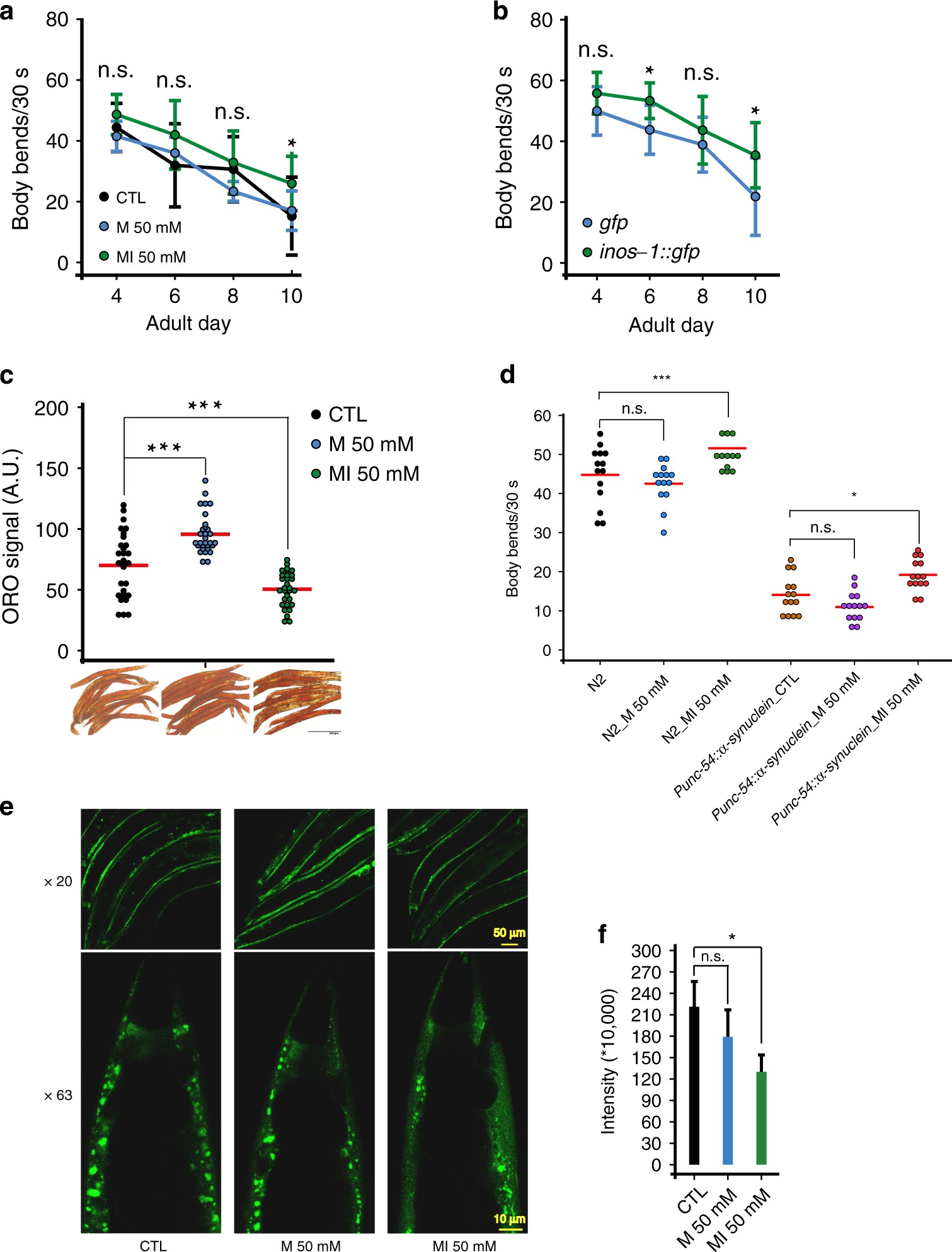
Fig. 2: MI supplementation alleviates worm health decline during aging.
a 50 mM MI alleviated worm mobility decline from AD_4 to AD_10 (Data are presented as mean values ± SEM, n = 10, two-sided t-test *p < 0.05 on AD_10), N = 2. b inos-1 OE alleviated worm mobility decline (Data are presented as mean values ± SEM, n = 10, two-sided t-test *p < 0.05 on AD_6 and AD_10), N = 2. c ORO staining shows MI suppressed worm fat storage on AD_10 (n = 30, two-sided t-test ***p < 0.001), N = 6. d MI promoted the mobility of the worm model of PD ([unc-54p::α-synuclein::YFP + unc-119(+)] (n = 14, two-sided t-test *p < 0.05), N = 3. e Representative images show MI supplementation decreased α-SYNNUCLEIN::YFP aggregates in this PD model (20× labels images under 20x microscopy, scale bar 50 μm; 63× for 63x microscopy scale bar 10 μm). f Quantification of Fig. 2e like worms but on one worm tail region per image, under 63x CONFOCAL microscopy (Data are presented as mean values ± SEM, n = 30, two-sided t-test *p < 0.05), N = 2. Source data are provided as a Source Data file.
MI was reported to reduce body fat in mammals33, and we found that MI also decreased worm fat storage after examining lipid oil red O (ORO) staining. In contrast, the osmotic control M increased worm body fat, compared to untreated control (***p < 0.001, Fig. 2c).
We also checked the effect of MI on worm reproduction and found that it did not obviously affect worm brood size when treating the worms from L4 stage to AD_8 (Supplementary Fig. 2c).
As MI supplementation enhanced worm mobility, we asked whether MI could rescue the phenotypes of worms modeling mitochondria-related disease. We found that MI but not M increased the mobility of Parkinson’s disease (PD) model (α-synuclein::YFP, *p < 0.05, Fig. 2d). Consistently, we found MI decreased the α-SYNUCLEIN puncta in the PD model (*p < 0.05, Fig. 2e, f).
MI promotes mouse healthspan
Next, we examined whether the health effects of MI were conserved in mouse. We treated middle-aged female C57BL/6J mice with 580 mg MI per kg bodyweight by gavage, twice per week, from 9 months of age to 12 months, we also set up CTL, Pre-treatment CTL and Young CTL groups, with the latter two groups from other batches of mice than CTL and MI. All mice were tested and sacrificed at the same time (Fig. 3a). For the mouse experiment, we chose the lowest MI concentration reported in literature33,34,35, 580 mg/kg. At this concentration, we did not observe any liver, kidney, or overall toxicity.
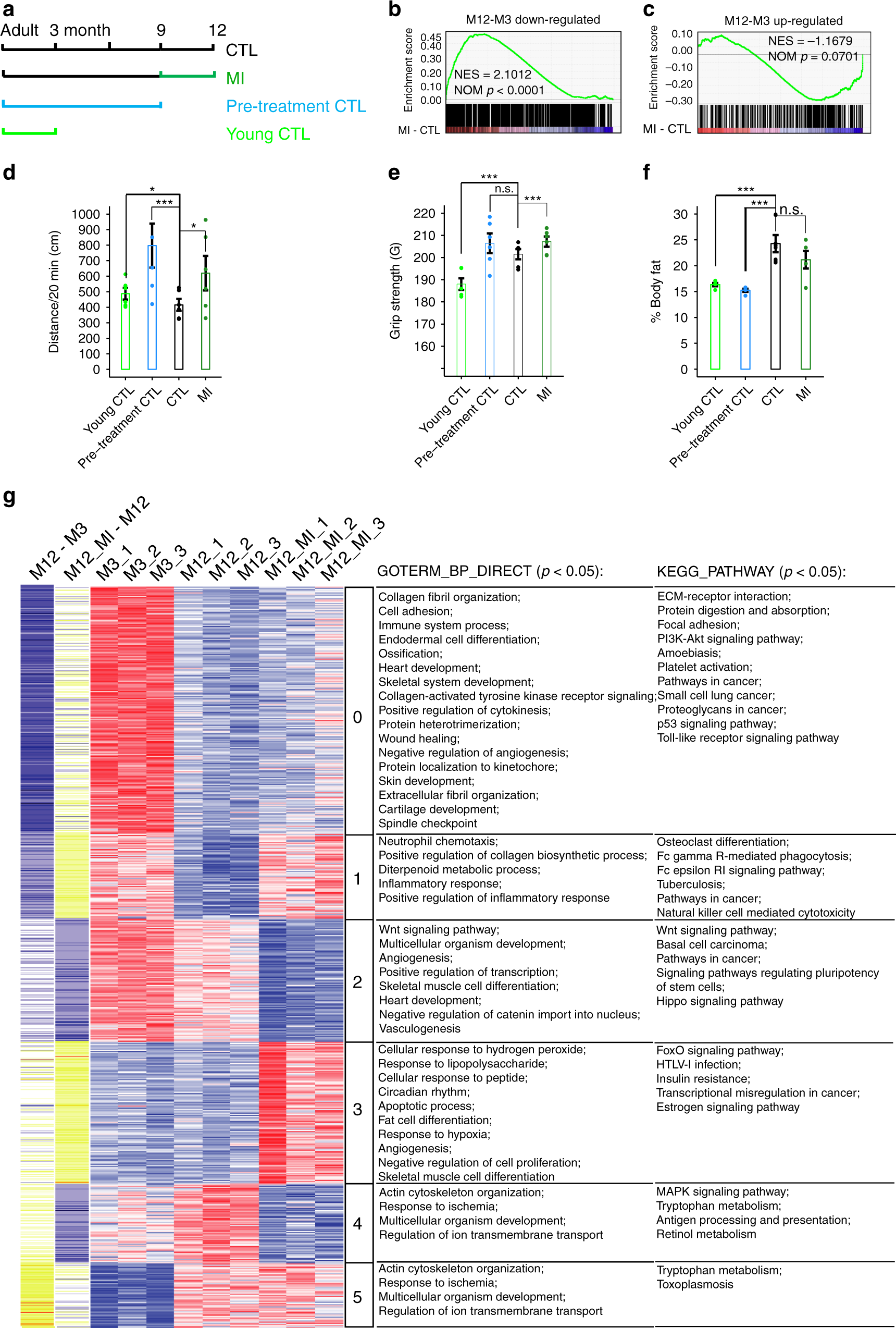
Fig. 3: MI promotes health in middle-aged mice.
a Design of mouse experiment. b, c GSEA results show mouse aging-downregulated or upregulated gene expressions, measured by RNA-seq log2-fold-change in FPKM, were reversely regulated by MI supplementary (GSEA p < 0.001 and p = 0.0701 respectively), N = 1. d MI increased mouse running distance in 20 min (Data are presented as mean values ± SEM, n = 6, two-sided t-test *p < 0.05, p = 0.028 between CTL and MI), N = 2. e MI increased mouse limbs grip strength (Data are presented as mean values ± SEM, n = 6, two-sided t-test *p < 0.05, p = 0.0001 between CTL and MI), N = 2. f MI decreased mouse body fat content (Data are presented as mean values ± SEM, n = 8 for CTL, n = 4 for others, two-sided t-test n.s. not significant, p = 0.12 between CTL and MI), N = 1. g Clustering of the aging-related genes, which were reversed by MI. Cluster 0 and 1 are aging downregulated genes, that were upregulated by MI, cluster 4 and 5 are aging upregulated genes, that were downregulated by MI. Cluster 2 or 3 show genes, which didn’t change obviously from M3 to M12, but were downregulated or upregulated by MI, respectively. Source data are provided as a Source Data file and Supplementary Dataset 1.
We first examined the transcriptome changes induced by MI in mouse muscle. MI treatment reversed the aging-related muscle transcriptome changes, as it did in the worm, i.e., it significantly increased the expression of aging downregulated genes (p < 0.0001, Fig. 3b), and showed a trend towards decreasing aging upregulated genes (p = 0.0701, Fig. 3c). The Pearson correlation coefficient (PCC) between changes induced by aging and by MI is −0.509, demonstrating a global anti-aging effect at the transcriptome level.
Phenotypically, the MI treated mice not only ran longer distance in open-field tests (*p < 0.05, ***p < 0.001, Fig. 3d), but also displayed more powerful muscle strength in grip-tests (***p < 0.001, Fig. 3e), MI was previously reported to decrease body fat33, and here we found MI induced a trend towards reduced body fat, although it was insignificant in this cohort of mice (p = 0.2, Fig. 3f).
Based on all expressed genes (FPKM > 0 in any sample), PCA of all nine samples (three biological replicates for all Young_CTL, CTL, and MI) revealed that MI treated muscle samples were closer to CTL samples on PC1 and similar to Young_CTL samples on PC2 (Supplementary Fig. 3a, PC1 = 46%, PC2 = 27%). Interestingly, the effect of MI mostly showed on PC2, which is positively related to mitochondrial function (Supplementary Fig. 3a–b). The most significantly upregulated genes by MI treatment, as compared to CTL samples, include Ddit4, Npas2, Arrdc3, Foxo1; the most downregulated genes include Cish, Zfp503 (Supplementary Fig. 3c).
We further examined the differentially expressed genes (FDR < 0.2) by clustering analysis. Cluster 0 and 1, related to cell adhesion and adaptive immunity, respectively, were downregulated genes comparing 12 vs 3-month mice, and were upregulated by MI; cluster 4 and 5 related to growth regulation and ion transport, respectively, were upregulated genes comparing 12 vs 3-month mice, and downregulated by MI. We also found MI induced downregulation of blood vessel development-related cluster 2 and upregulation of transcription related to cluster 3 genes, which were not drastically changed from M3 to M12 (Fig. 3g).
The MI derivative PI(4,5)P2 is the effector for MI induced longevity
Next, we asked whether MI exerts its effects through its derivatives in the metabolic pathway. To address this, we examined the lifespan of worms with LOF of enzymes in all steps of the MI metabolic pathway, by using RNAi or mutants (Fig. 4a; Supplementary Fig. 4a–o, Supplementary Fig. 5, Supplementary Table 3). First, we looked into the inositol pyrophosphates of the MI derivatives and found that reduction of IP8 by knocking down its synthases, F46F11.1 (step 27) and F30A10.3 (step 28), decreased worm lifespan (p < 0.02, Supplementary Fig. 4m and p = 0.08, Supplementary Fig. 4n, respectively). IP7 was reported to be an Akt inhibitor in mouse28, IP6 supplementation also extended worm lifespan in a dose-dependent manner (Supplementary Fig. 4p). Worms cannot produce IP7 from MI, due to the lack of an enzyme to synthesize IP6 from IP5 (Fig. 4a; Supplementary Fig. 1a, b). Therefore, MI’s longevity effect is unlikely to be attributed to IP7.
.../...
.
Edited by Engadin, 16 September 2020 - 03:24 PM.















