































.
F U L L T E X T S O U R C E : Portland Press
Abstract
Among several theories to explain the complicated process of human ageing, the mitochondrial oxidative stress hypothesis has received recent attention. Considering that lifespan and ageing rates vary considerably across taxa, a better understanding of factors that lead to negligible or extremely rapid senescence in mammals may generate novel approaches to target human ageing. Several species, such as naked mole rats, ocean quahog, rockfish and Greenland shark, have been identified that exhibit negligible senescence and superior resistance to age-related diseases. Considering that the available literature suggests that their outstanding stress resistance is linked to maintenance of protein homeostasis and robust mitochondrial functions, treatments that target protein modification and upregulation of matrix antioxidants may have implications for extending human health span.
Introduction
The principles of ageing and an inescapable advancement towards death have always captivated the interest of mankind. Today the hunt for the ‘fountain of youth' has been replaced by the search for geroprotectors; i.e. pharmacological or nutraceutical compounds that prevent ageing [1]. Though life and health spans have been prolonged in many experimental animals, it is arduous to conduct studies to test if these regimens also prolong life and health span in humans [2]. As the main cause of accelerated mammalian ageing is energy excess, caloric restriction without malnutrition is currently the most effective non-genetic intervention to delay ageing phenotypes in a wide range of mammals from flies [3] to primates [4]. A problem with this strategy is that caloric restriction may have adverse effects [4] and reduce the quality of life in humans. An alternative approach to arrest ageing would be to use drugs that have caloric restriction mimetic effects [5]. Sirtuins are believed to mediate the pro-longevity effect of calorie restriction; thus, sirtuin activators have attracted interest within gerontology. As the human ageing process is slow and gradual, clinical trials of anti-senescence drugs [6], nuclear factor erythroid 2-related factor 2 (NRF2)-agonists [7], NAD-dependent deacetylase sirtuin-1 (SIRT1) agonists [8] and/or dietary interventions that prolong lifespan in animals, such as caloric [4] and methionine [9] restriction, would need to be extremely long.
Considering that the global number of individuals >80 years old will triple by 2050, ageing is a considerable challenge for biomedical research that will have a dramatic influence on human morbidity and mortality [10]. Diseases of ageing are influenced by mechanisms that show strong evolutionary conservation and ageing is the main risk factor for a number of ‘burden of life style' diseases, such as cancer, cardiovascular disease (CVD), obesity, type-2 diabetes, osteoporosis, Alzheimer and chronic kidney disease (CKD), that have become exceedingly prevalent in the modern society [7]. Detailed studies of relatively large groups of patients displaying features of accelerated ageing, such as CKD, may be an opportunity to learn more about slow ageing processes [11]. The majority of chronic diseases that accumulate with age share common underpinning mechanisms, such as low-grade inflammation, oxidative stress, mitochondrial dysfunction, metabolic imbalances and senescence [12]. The role of cellular senescence in diseases that accumulate with age is a growing topic in gerontology; cellular senescence being integral to the physiology of ageing, longevity and diseases that accumulate with age, either by protein–protein interactions, allocation of common genes or prevailing signalling pathways [13]. A major problem in modern health care is the growing gap between health span and lifespan; a gap that has major economic and societal implications. Thus, rather than focus on increasing lifespan, gerontology research should aim to increase health span. Strategies that target central ageing structures in the whole cluster of ‘burden of life style' diseases would likely reduce the gap between health span and lifespan [7].
Studies of longevity in the animal kingdom—could it provide clues for human ageing?
Studies of long-lived animals that stay healthy and capable of reproduction until death (i.e. long health span) could provide clues on how to target central human ageing processes [7]. Although the lifespan trajectories in different model organisms depict remarkable similarities in the ageing process with humans [14], many wild animals may not have the time to develop features of senescence, as they die of disease, are predated or starve before they reach sufficiently advanced years [15,16]. In order to activate extra resources for survival during stressful periods of the animals life, such as hibernation, it has been speculated that ageing programmes are dampened during such periods [17].
One of the biggest challenges in gerontology is to identify changes in cells that are related to senescence (cell cycle arrest with loss of a cells ability of division and growth), instead of just chronology [18]. Senescence has been considered trivial in wild animals. However, current evidence implies that it is widespread in natural populations of mammals and birds, affecting survival and reproduction of older individuals. A problem with existing data on longevity in animals is that they often consist of maximum values for lifespan (usually single observations) rather than sound and statistically robust estimates of their mean longevity [19]. Delineating the physiological and biochemical causes of ageing process in animals is a developing area of research for several reasons. These include the possible assets for human well-being, the relation of ageing to growth and reproduction and the responses of organisms to natural conditions and stress.
From our archaic past, when our common nematode-like ancestors aged exceptionally rapidly, mutations have expanded human lifespan more than 2000-fold [20]. Given that protracted longevity in animals is associated with augmented stress resistance, longevity requires uninterrupted and large-scale alterations of organs to suspend senescence. Studies of animals with particularly long lifespans would be an approach to better understand mechanisms linked to superior functionality and learn about ageing processes and find central targets for extending health span [21]. A recent study that included a variety of birds and mammals with different lifespans and body sizes showed that telomere shortening rate predicts lifespan in animals [22]. In the following, we discuss what we can learn from animal species that during evolution have adapted systems that render them extraordinary long-lived for their size (Figure 1).
Figure 1.
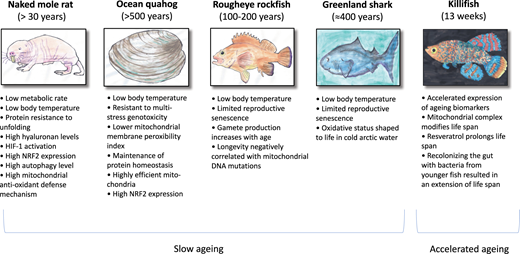
Animals of interest in ageing research: Reported characteristics of four long-lived animals (naked mole rat, ocean quahog, rougheye rockfish, Greenland shark) with negligible senescence and one animal showing accelerated ageing (turquoise killifish).
Naked mole rats
Among long-lived animals, none has received as much recent attention as the naked mole rat (Heterocephalus glaber), a native of Eastern Africa that has emerged as a model organism for studies of ageing-related diseases and negligible senescence [23]. In contrast to other mammals, naked mole rats do not conform to the Gompertzian law of age-related mortality risk [24]; i.e. their risk of death does not increase with age. During evolution, these rodents have adapted to severe constant hypoxia in underground burrows with limited gas exchange. They can survive up to 18 min of anoxia without apparent injury, due to fructose-driven glycolysis [25]. It has been reported that hypoxia-inducible factors (HIF)-1α activation is another important mechanism for their remarkable hypoxia adaption [26]. Naked mole rats live >5 times longer than similarly sized rodents and >9 times longer than laboratory rats and show sustained good health during their lifetime. Notably, naked mole rats are subjected to high levels of oxidative stress and show increased levels of oxidative damage and less robust repair mechanisms than their short-lived rodent counter-parts, but still maintain a superior health span; this is not consistent with the free radical theory of ageing [27]. Beside superior anoxia tolerance, they are resistant to age-related diseases, such as neurodegeneration, CVD and cancer [21]. Out of 1000's of necropsies only two cases of cancer have been reported in naked mole rats [28]. In addition, naked mole rats experience no changes in body composition, or decline in genomic and proteomic integrity due to elevated proteasome quality control mechanisms [29]. Their diet (well-off in tubers and roots) provide not only a rich source of anti-inflammatory and anti-oxidative polyphenols, but is also restricted in calories and methionine. It is likely that their diet in combination with low body temperature in a hypoxic and hypercapnic milieu has resulted in a series of metabolic adaptions that contribute to their exceptional resilience against age-associated diseases. Efficient protein stability and integrity with less urea-induced protein unfolding, less protein ubiquitination and higher proteasome activity [30], high levels of autophagy [31], high hyaluronan skin expression [32], a superior capacity for mitochondria to consume H2O2 [33] and overexpression of NRF2 [23] are reported factors liable for maintaining superior damage control mechanisms (Figure 1).
Secrets of longevity found in oceans
Fish (the most diverse class of vertebrates with >20 000 existing species) provide an enormous spectrum of longevity that could be used for gerontology research. Although some marine animals, such as sea turtles, sharks and sturgeons are reported to have a long lifespan, they have not been extensively used for studies of ageing. In contrast to the common contention that turtles (Chrysemys picta) do not age, data spanning >20 years from a wild population of turtles has shown that survival declines among adults [34]. Fish exhibit three types of senescence; rapid (such as eels, killi fish and pacific salmon), gradual (such as guppy and platyfish) and negligible (such as sturgeons and rockfish). Given recent data indicating that senolytic drugs, such as quercetin and navitoclax, may be an option to arrest human ageing processes [35] comparative studies of fish with rapid or negligible senescence would be of interest [36]. Whereas lower environmental temperature and dietary restriction retards the ageing process in species showing gradual senescence, the reason(s) why only some species of fish exhibit negligible senescence are undisclosed [36]. The existence of fish with negligible senescence allow extensive studies of cellular, genetic and biochemical characteristics of slow ageing. It has been reported that muscle extracts from fish with negligible senescence, such as sturgeon, have protective effects on senescence induced by oxidative stress [37]. It would also be interested to find out if long-lived fish exhibit an enhanced capacity to repair DNA damage, protect protein function or exhibit a continued turnover of tissue. The rockfish is a long-lived fish with a documented age range of 100–200 years, that has attracted interest as a model system due to intermediate growth (i.e. it continues to increase in body size throughout life) and limited reproductive senescence. In contrast to other mammals, gamete production in female rockfish increases with age and size fecundity is positively correlated with age [38]. Among long-lived fish, sharks have received recent attention since they live much longer than realized previously [39]. Radiocarbon studies of the eye lens in the Greenland shark (Somniosus microcephalus) show that they may reach an age of almost 400 years; i.e. they are the longest-lived vertebrate documented [40] and surpass the maximum age of >200 years reported in bowhead whales (Balaena mysticetus) [41].
The most long-lived marine species are bivalve molluscs, such as the ocean quahog, which provide the widest range of lifespans available for interspecies comparisons. The maximal lifespan in different populations of bivalve molluscs (Spisula solidissima vs. Arctica Icelandica) ranges between 36 to >500 years (age is counted by growth rings on the shell) despite the similar size and living conditions [42]. Thus, Arctica Icelandica is the longest-lived non-colonial metazoan known on the planet. As bivalves accommodate a protracted sphere of environmental conditions and express markedly contrasting metabolic phenotypes they constitute a well-founded model to study ageing processes [42]. Their exceptional longevity appears independent of telomere length regulation [43]. Moreover, whereas their longevity-related differences in peroxidation index seem resilient to environmental conditions, such as temperature, season, nutrition and age [44] they are resistant to multi-stress genotoxicity [45]. To elucidate the underpinning features of successful cardiac ageing, age-related changes in proteosome activity, inflammatory markers, heat shock proteins and mitochondrial complexes were analysed in the heart of A. Icelandica [46]. The results showed that maintenance of protein homeostasis preserves cardiac function and that levels of oxidatively damaged proteins did not change over the first 120 years of their life [46]. Treaster et al. [47] have also emphasized the relationship between proteostasis and longevity through estimation of function, unfolding and aggregation. As inhibition of Hsp90 protects from atherosclerosis via cytoprotective mechanisms that are NRF2-dependent [48] and SKN-1/NRF2 is essential for the health span gains provided by metformin treatment in C. elegans [49], a role for NRF2 as a pro-longevity signalling pathway across phyla is supported. Indeed, a transcriptomics-based screening recently identified that inhibition of Hsp90 deferred ageing by augmented protein homeostasis in C. elegans [1].
Free radical and mitochondrial theories of ageing
According to the free radical theory of ageing (reactive oxygen species [ROS] arise as bi-products of metabolism), long-lived species should be more resistant to oxidative damage and senescence than short-lived species. However, as research using 18 different genetic manipulations of the antioxidant defense system showed no effect on lifespan in mice (except deletion of the Sod1 gene), the evidence argues against the hypothesis that oxidative damage plays a dominant role in ageing [50]. Data from animal species other than mice have shown equivocal results. It was reported that augmented resistance to oxidative stress and maintenance of protein homeostasis preserves cardiac function in A. Icelandica [46] and may be a prerequisite for their extreme longevity [51]. When differences in markers of oxidative stress were compared in mollusc species with a six-fold difference in lifespan, Gruber et al. [51] reported correlations between DNA oxidation and biological, but not chronological age, implying that nucleic acid oxidation reflected intrinsic ageing mechanisms. On the other hand, since surrogate markers of oxidative damage in red blood cells and muscle did not correlate to lifespan in Greenland sharks, the free radical theory of ageing do not apply to all long-lived species [52]. In the case of the Greenland shark, the oxidative status may rather be shaped to the life in cold and deep artic water [52].
It has been suggested that the cumulative effect of biochemical damage in mitochondria causing mitochondrial mutations plays a key role in many chronic diseases associated with premature ageing, such as CKD, type-2 diabetes and neurodegenerative diseases. The transcriptional coactivator peroxisome proliferator-activated receptor gamma co-activator 1 (PGC1α), the master regulator of mitochondrial biogenesis, allow animals to conform their energy metabolism to their current state of nutrition and their environment. It is likely that the evolution of differences in the distribution and density of mitochondria play a major role for longevity, aerobic capacity and protection in extreme conditions. In the animal kingdom, the levels of mitochondrial enzymes vary more than 100-fold, from fish white muscle to the flight muscles of birds [53]. The Atlantic bluefin tuna (Thunnus thynnus) is a migratory fish with a remarkable swimming endurance and high metabolic demands, which stands out from other fish because they possess red muscle (due to an exceptionally high content of mitochondria. In a recent study, Dumesic et al. [54] demonstrated that the upstream open reading frame (uORF), which slows down the production of PGC1α, was completely absent in the Atlantic blue fish tuna. Their finding indicates that disruption of uORF may be an important evolutionary adaption in the animal kingdom that allows high metabolic demand.
If the rate of mitochondrial mutation is negatively correlated with longevity, the accumulation of mitochondrial mutations should associate with the degree of senescence [13]. Indeed, Blier et al. [42] reported lower ROS production and a lower mitochondrial membrane peroxibility index in the long-lived A. Icelandica, supporting the concept that mitochondrial dysfunction-mediated dysregulation of molecular and metabolic pathways boosts senescence. Considering that A. Icelandica produces markedly lower rates of H2O2 compared to short-lived species and carries mitochondria that are remarkably efficient in energy transduction, extreme longevity seems to be associated with robust and efficient mitochondria that produce low levels of H2O2 and ROS [42]. Given the reported links between mitochondrial dysfunction and cellular senescence [55], and recent data showing that NRF2 activation plays a role in mitochondrial quality control [56], it is not surprising that the NRF2 signalling pathway plays an important role in the fine-tuning of gene expression in bivalve molluscs [57]. In accordance, a superior antioxidant mitochondrial defence system [33] together with overexpression of NRF2 [23] characterizes the exceptionally long-lived naked mole rat. The evolvement of kelch-like ECH-associated protein 1 (KEAP1) provided mammals with a more precise way to regulate NRF2 activity [58]. Considering that the lifespan of rockfish (Sebastes) is negatively correlated with the rate of mitochondrial DNA mutations [38], this further support the concept that mitochondria play a main role for longevity in the animal kingdom. Data in birds and mammals corroborate these findings and show an inverse relationship between longevity and evolutionary rate of mitochondrial proteins [59]. A comparative study in 21 bird species showed that red blood cells from long-lived birds produced less mitochondrial ROS than red blood cells of short-lived birds [60]. However, although the majority of studies report that a reduced lifespan in mammals is related to an increase in somatic mitochondrial mutations [61,62], Vermulst et al. [63] reported that mitochondrial point mutations do not limit the natural lifespan of mice. In accordance, a study in C. elegans reported that mitochondrial DNA damage does not control lifespan [64].
.../...
F O R T H E R E S T O F T H E S T U D Y, P L E A S E V I S I T T H E S O U R C E .
.














