































The protein-import systems of organelles can become clogged by proteins. A protein from one organelle, the endoplasmic reticulum, is found to also unclog such blockages in mitochondrial organelles.
Organelles known as mitochondria are the energy-generating powerhouses of the cell. As a crucial part of the cellular machinery, disruption to their function could have serious consequences, and so mechanisms exist to combat mitochondrial dysfunction. Writing in Nature, Mårtensson et al.1 report a pathway that tackles problems concerning the import of proteins into mitochondria, as they arise, to prevent mitochondrial and cellular dysfunction. This pathway uses part of the cellular repair kit that handles damaged proteins in another organelle — the endoplasmic reticulum (ER).
At least 1,000 proteins reside within the mitochondria of yeast, where they carry out functions that contribute to cellular energy production and metabolism2. Almost all of these proteins are made in the cytoplasm and are then transported into mitochondria in an unfolded precursor form. Mitochondria receive a heavy inbound flow of protein traffic, and the entry route that bears the brunt of this influx is a protein complex on the mitochondrial surface called the TOM complex. When the TOM complex becomes overloaded or is compromised due to mitochondrial damage, unfolded mitochondrial proteins can accumulate in the cytoplasm and cause cellular toxicity3–6. Mechanisms exist to combat the stress to cells that results from this3–6, but how mitochondria handle protein-import failure as it arises was unclear.
The pore of a TOM complex will sometimes become clogged with a mitochondrial protein (Fig. 1). Protein misfolding is a common cellular event that can be exacerbated by stress or occur if a protein is in a mutant form7. A mitochondrial precursor protein that folds prematurely in the pore of the TOM complex could become stuck there. A blockage might also occur if the energy levels of the cell drop, leading to a slowing or stalling of protein import8. If such an obstruction is not removed, the accumulation of unfolded precursor proteins in the cytoplasm can trigger transcriptional changes that lead to the unleashing of a cellular stress response4. Such stress-response pathways3,4 include the UPRam pathway and the mitoCPR pathway, which prime the cytoplasmic waste-disposal system — a protein complex called the proteasome — to degrade the accumulated precursor proteins.
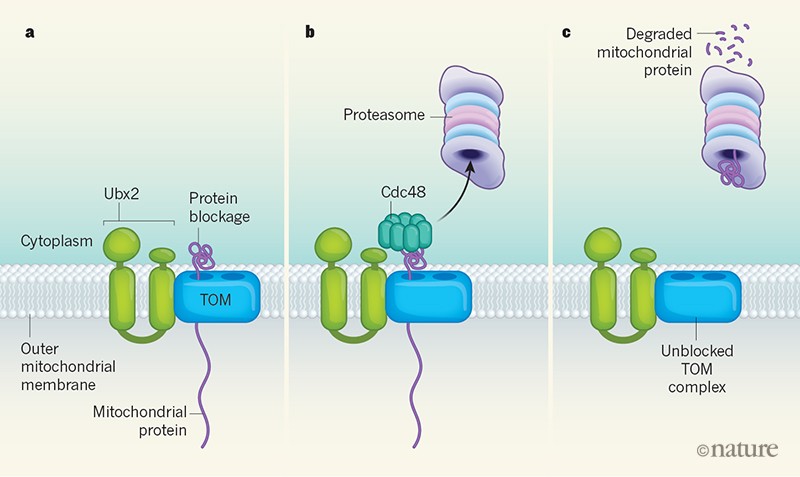
Figure 1 | A mechanism for unclogging blockages in the protein-import system of mitochondria. a, Unfolded mitochondrial proteins are imported from the cytoplasm into mitochondria through a protein complex called TOM that is located on the outer mitochondrial membrane. However, the TOM complex can become blocked when, for example, a protein folds prematurely as it passes through the complex. Mårtensson et al.1 report a system in yeast cells that monitors and unclogs such blockages. The protein Ubx2, which acts in quality-control systems for another organelle (the endoplasmic reticulum)10,11, is required to recognize protein blockages in the TOM complex. b, Ubx2 recruits the protein Cdc48, which removes the clogged protein and delivers it to a protein-degradation complex called the proteasome. c, The degradation of the mitochondrial protein completes the process that unclogs the TOM complex. Figure based on Fig. 4f of ref. 1.
Mårtensson et al. investigated whether a system is in place to constantly monitor the TOM complex and to remove trapped precursor proteins, thereby preventing blockages from hampering protein import to an extent that causes cellular stress. Such a monitoring system is in place for an entry gate into the ER9. To identify possible factors that might clear away trapped precursor proteins, Mårtensson et al. purified the TOM complex from the mitochondria of yeast cells. One of the TOM-complex-associated proteins that they identified was Ubx2, which was a surprise. This is because Ubx2 is best known for its activity at the ER, where it functions in the routine clearance of misfolded proteins as part of an ER-specific quality-control pathway called ER-associated degradation (ERAD)10,11. Mårtensson and colleagues conducted biochemical experiments that showed that Ubx2 exists in two distinct cellular pools, one at the ER and another on the mitochondrial surface that is associated with TOM complexes.
In the ERAD pathway, enzymes add the protein ubiquitin to misfolded proteins that are trapped in the ER10,11. This ubiquitin tag is recognized by Ubx2, which triggers the recruitment of a protein called Cdc48 — a type of enzyme called an AAA ATPase. Cdc48 extracts misfolded proteins from the ER and delivers them to the proteasome for degradation10,11. To ascertain whether Ubx2 might function in mitochondria in a similar way, Mårtensson and colleagues tested whether Cdc48 interacts with the TOM complex. They found that it does, and also that the interaction depends on Ubx2. When the authors blocked the energy source that is needed for mitochondrial protein import, this resulted in an accumulation of precursor proteins only in mitochondria that lacked Ubx2.
Precursor-protein accumulation in the absence of Ubx2 was also observed if mitochondrial protein import was slowed by an alternative means — the use of a mutant version of a protein that drives protein import into mitochondria12,13. In this set-up, Mårtensson et al. observed that precursor proteins accumulated in the cytoplasm of cells that had defects in Cdc48 or in a component of the proteasome, which indicates that Cdc48 and the proteasome have a key role in this pathway.
To specifically determine whether Ubx2 is needed to remove precursor proteins that are clogging the TOM complex, Mårtensson and colleagues expressed mitochondrial precursor proteins, in yeast, that were chosen because they are imported slowly into the organelle. The authors engineered this scenario to boost the number of blockage events, and then extracted TOM complexes. They found that, in cells that lacked Ubx2, precursor proteins were stuck in TOM complexes. However, in cells where Ubx2 was present, no precursor proteins were detected in TOM complexes. This key piece of evidence indicates that Ubx2 is part of a previously unknown quality-control mechanism that surveils the TOM complex, and Mårtensson et al. have named it the mitochondrial protein translocation-associated degradation (mitoTAD) pathway.
Mårtensson and colleagues’ findings raise two crucial questions. First, what is the relationship between the mitoTAD pathway and the mitochondrial quality-control pathways that operate under conditions of cellular stress? During their investigation, the authors unearthed a functional link between Ubx2 and the protein Vms1, as well as with the enzyme Msp1 (which, like Cdc48, is an AAA ATPase). Both Vms1 and Msp1 have been implicated in mitochondrial quality-control processes14–17. When the authors removed these proteins from cells that lacked Ubx2, mitochondria lost their energy-producing function and ubiquitin-tagged proteins accumulated. This result suggests that Vms1 and Msp1 work in a way that is complementary to Ubx2 function. An investigation to assess the crosstalk between these pathways and the mitoTAD pathway might deepen our understanding of mitochondrial quality-control mechanisms.
Second, how does Ubx2 recognize that a precursor protein is stuck in the TOM complex? Ubx2 can bind to ubiquitin, so ubiquitin is probably used to tag clogged precursor proteins. Defining the components of the ubiquitination machinery and the ubiquitin signals that govern not just the mitoTAD pathway, but also the stress-induced quality-control pathways that regulate mitochondrial protein import will be an important direction for future research. The fact that the ER and mitochondria use the same protein for quality control might imply that these processes are co-regulated.
Many other fascinating questions also remain to be addressed. In the meantime, Mårtensson and colleagues’ work has painted an illuminating picture of a previously unknown aspect of how mitochondria protect their main entry gate.















